
Abstract
The natural infection of sand flies by Leishmania species was studied in the Andean areas of Peru where cutaneous leishmaniasis caused by Leishmania (Viannia) peruviana is endemic. Sand flies were captured by human bait and Center for Disease Control (CDC) light trap catches at Nambuque and Padregual, Department of La Libertad, Peru, and morphologically identified. Among 377 female sand flies dissected, the two dominant man-biting species were Lutzomyia (Helcocyrtomyia) peruensis (211 flies) and Lutzomyia (Helcocyrtomyia) caballeroi (151 flies). Another sand fly species captured by light trap was Warileya phlebotomanica (15 flies). The natural infection of sand flies by flagellates was detected in 1.4% of Lu. (H.) peruensis and 2.6% of Lu. (H.) caballeroi, and the parasite species were identified as Le. (V.) peruviana and Trypanosoma avium, respectively, by molecular biological methods. The results indicated that the vector species responsible for the transmission of leishmaniasis in the study areas is Lu. (H.) peruensis. In addition, the presence of Trypanosoma in man-biting sand fly species means that more careful consideration is necessary for vector research in areas of Andean Peru where leishmaniasis is endemic.
Introduction
Cutaneous leishmaniasis (CL), commonly known as Uta, mostly caused by Leishmania (Viannia) peruviana, is highly endemic in the Andean areas of Peru (Lucas et al. 1998, Davies et al. 2000, Arevalo et al. 2007, Kato et al. 2010c). To date, Lutzomyia (Helcocyrtomyia) peruensis (Perez et al. 1994, 2007, Kato et al. 2008) and Lutzomyia (Helcocyrtomyia) ayacuchensis (Caceres et al. 2004) are proven vectors of Le. (V.) peruviana in the Peruvian Andes, and Lutzomyia verrucarum (Verrucarum group) has exhibited a capacity for transmission under experimental conditions (Davies et al. 1993). Flagellates have been also observed in the gut of Lutzomyia (Helcocyrtomyia) noguchii, a species that prefers to feed on field mice, in an Andean highland area of Peru (Caceres et al. 2004). Although not characterized, the parasite tested negative in a polymerase chain reaction (PCR) specifically targeting the subgenus Leishmania (Viannia), suggesting the prevalence of flagellates other than Le. (V.) peruviana in the Peruvian Andes. To date, nonpathogenic flagellates such as Endotrypanum species, an intraerythrocytic parasite of sloths, and other trypanosomatids such as Crithidia, Blastocrithidia, Herpetomonas, and Phytomonas have been detected within sand fly species in tropical and subtropical areas of the New World (Shaw 1964, Kato et al. 2007, Rocha et al. 2010). In addition, Trypanosoma species of a mammal, lizards, snakes, and toads were reported to be transmitted by sand fly species in tropical, subtropical, and desert areas of the African, American, and recently Asian continents (Anderson and Ayala 1968, Ayala 1971, Ayala and McKay 1971, Naiff et al. 1989, Ferreira et al. 2008, Lemos et al. 2008, Viola et al. 2008, Kato et al. 2010b). As the flagellated forms of these parasites in the insect gut are morphologically similar to those of Leishmania, careful differentiation is needed for the epidemiological study of the vectors responsible for the transmission of Leishmania species.
In the present study, to further define circulating vector species and natural infections by Leishmania, sand flies were investigated in the Otuzco Province, Department of La Libertad located in the Andean highlands of the northern Peru where CL caused by Le. (V.) peruviana is endemic (Kato et al. 2010c).
Materials and Methods
Sand fly collection
The natural infection of sand flies by Leishmania was examined in the Andean areas of the northern Peru where CL caused by Le. (Lv.) peruviana is endemic (Kato et al. 2010c). Sand flies were captured using protected human bait on the 17–20th of August 2008 and CDC light traps on the 11–13th of January 2009 at Nambuque (2300 m above sea level) and Padregual (2300 m above sea level), both neighboring areas of La Cuesta (7° 54′S, 78° 42′W) in the Province of Otuzco, Department of La Libertad. All the flies were captured between 19:00 p.m. and 20:30 p.m. by protected human bait and between 18:00 p.m. and the next 6:00 a.m. by CDC light traps. The captured sand flies were dissected and the species were identified mainly based on the morphology of their spermathecae (Young and Duncan 1994). These flies were also microscopically examined for Leishmania in the gut at 400 × magnification. The flagellates in the gut were fixed in absolute ethanol and stored at room temperature for molecular biological analyses. In some cases, the flagellates were inoculated into Difco blood agar biphasic medium containing 20% defibrinated rabbit blood and later cocultured with Spodoptera frugiperda Sf9 insect cells in Grace's medium supplemented with 10% fetal calf serum (Kato et al. 2010b). In the study areas, 10 cases suspected for CL were found during the research periods, and infection of Le. (V.) peruviana was molecular biologically identified in four cases.
Identification of flagellate species
The ethanol-fixed specimens were individually lysed in 50 μL of DNA extraction buffer (150 mM NaCl, 10 mM Tris-HCl [pH 8.0], 10 mM ethylenediaminetetraacetic acid, and 0.1% sodium dodecyl sulfate) in the presence of proteinase K (200 μg/mL), and 0.5 μL of the extract was directly used as PCR templates (Kato et al. 2007, 2008). The cytochrome b (cyt b) gene was amplified from the flagellates with primers prepared for the leishmanial cyt b gene (L.cyt-S: GGTGTAGGTTTTAGTYTAGG and L.cyt-R: CTACAATAAACAAATCATAATATRCAATT) using Ampdirect Plus reagent (Shimadzu Biotech, Tsukuba, Japan) (Kato et al. 2007, 2008).
For the identification of Trypanosoma species, the glycosomal glyceraldehyde phosphate dehydrogenase (gGAPDH) and small subunit ribosomal RNA (SSU rRNA) genes were also amplified from the flagellate using primers specific for the gGAPDH gene (G1: CGCGGATCCASGGYCTYMTCGGBAMKGAGAT and G4a: GTTYTGCAGSGTCGCCTTGG) and the SSU rRNA gene (TRY927F: GAAACAAGAAACACGGGAG and TRY927R: CTACTGGGCAGCTTGGA) of trypanosomatids (Noyes et al. 1999, Hamilton et al. 2004). The products were cloned into the pGEM-T Easy Vector System (Promega, Madison, WI) and sequences were determined by the dideoxy chain termination method using a BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA).
Le. (V.) peruviana was differentiated from Leishmania (Viannia) braziliensis by a PCR–restriction fragment length polymorphism (RFLP) analysis of the mannose phosphate isomerase (MPI) gene as described previously (Kato et al. 2008).
Phylogenetic analysis
The cyt b gene sequences were aligned with Clustal W software (Thompson et al. 1994) and examined using the program MEGA (Molecular Evolutionary Genetics Analysis) version 4.0 (Tamura et al. 2007). Phylogenetic trees were constructed by the neighbor-joining method with the distance algorithms in the MEGA package. Bootstrap values were determined with 1000 replicates of the datasets. The database for phylogenetic analyses consisted of cyt b gene sequences from Leishmania (Leishmania) infantum (GenBank accession number: AB095958), Le. (Le.) donovani (AB095957), Le. (Le.) major (AB095961), Le. (Le.) tropica (AB095960), Le. (Le.) amazonensis (AB095964), Le. (Le.) mexicana (AB095963), Le. (V.) panamensis (AB095968), Le. (V.) guyanensis (AB095969), Le. (V.) braziliensis (AB095966), Le. (V.) peruviana (AB433282), Le. (V.) lainsoni (AB433280), Le. (V.) naiffi (AB433279), and Le. (V.) shawi (AB433281), and Endotrypanum species, E. schaudinni and E. monterogeii (Uezato et al. Unpublished). The gGAPDH gene sequences were analyzed with those of Trypanosoma avium (FJ649487, AJ620263), T. brucei (XM 840453), T. congolense (AJ620290, AJ620291), T. corvi (FJ649496), T. cruzi (AJ620270, XM807045), T. grosi (AB362558), T. kuseli (AB362413), T. lewisi (AJ620272), T. microti (AJ620273), T. mega (AJ620253), T. rangeli (AF053742), and Trypanosoma species isolated from a sand fly (AB520637). The SSU rRNA gene sequences were analyzed with those of T. avium (AF416563, AJ009140, AY099319, AF416559), T. brucei rhodesiense (AJ009142), T. congolense (AJ009145, AJ009146), T. corvi (AY461665), T. cruzi (AJ009149, AJ009150), T. kuseli (AB175626), T. mega (AJ009157), T. microti (AJ009158), T. rangeli (AJ012417), and Trypanosoma species isolated from sand flies (AB520638, EU021237, EU021241, EU021242, EU021243, EU021244, EU021245), mosquitoes (AF416561), and hippoboscid flies (AF416562).
Results
In the present study, 377 female sand flies were dissected for identification at the species level, and three species of the genus Lutzomyia were recognized. Among them, the two dominant species were identified as Lu. (H.) peruensis (211 flies) and Lutzomyia (Helcocyrtomyia) caballeroi (151 flies), both of which are man-biting species. Another sand fly species captured by CDC light trap was Warileya phlebotomanica (15 flies). The natural infection of sand flies by flagellates was detected mainly in the midgut and hindgut of three Lu. (H.) peruensis and in the hindgut of four Lu. (H.) caballeroi (Table 1). Flagellate cyt b gene fragments were successfully obtained from six [three Lu. (H.) peruensis and three Lu. (H.) caballeroi] of the seven positive sand flies, and the nucleotide sequences were analyzed. The cyt b gene sequences from the three Lu. (H.) peruensis (08-per1, 08-per2, and 09-per) (GenBank accession numbers: AB566380, AB566381 and AB566382, respectively) had a greater degree of homology with those of Le. (V.) peruviana and Le. (V.) braziliensis (99.5%–99.9%) than the sequences of other Leishmania species (88.4%–98.8%). On the other hand, the cyt b gene sequences from the three Lu. (H.) caballeroi (08-cab3, 08-cab4, and 09-cab) (GenBank accession numbers: AB566377, AB566378, and AB566379, respectively) showed only 75.1%–77.1% homology with those of Leishmania species and 77.6%–77.7% homology with those of Endotrypanum species, which are flagellate parasites of mammals transmitted by sand flies (Shaw 1964, Kato et al. 2007). A phylogenetic tree showed that the three samples from Lu. (H.) peruensis divided into the clade of Le. (V.) peruviana and Le. (V.) braziliensis, whereas the other three from Lu. (H.) caballeroi were located far from both Leishmania and Endotrypanum species (Fig. 1). The three flagellates from Lu. (H.) peruensis were further differentiated by a PCR-RFLP–based analysis of the MPI gene targeting a single-nucleotide polymorphism (Kato et al. 2008). The RFLP pattern of all the three samples corresponded to that of Le. (V.) peruviana but not Le. (V.) braziliensis (data not shown), indicating that the flagellates within Lu. (H.) peruensis were Le. (V.) peruviana. As the flagellates from Lu. (H.) caballeroi were identified as neither Leishmania nor Endotrypanum species based on the cyt b gene analysis, the sequences were analyzed with the BLASTn program. The sequences showed 81% identity with those of T. cruzi and T. dionisii, suggesting that the flagellate belongs to the genus Trypanosoma. For further characterization, the gGAPDH and SSU rRNA genes of the flagellates were analyzed because these genes have been well studied in trypanosomatids (Noyes et al. 1999, Hamilton et al. 2004). The gGAPDH and SSU rRNA genes of the parasite 09-cab (GenBank accession numbers: AB566383 and AB566384, respectively) had >98% and 99% sequence identity, respectively, with those of T. avium. The SSU rRNA genes were also analyzed in parasites 08-cab3 and 08-cab4 (GenBank accession numbers: AB76100 and AB76101, respectively) and found to have >99% identity with that of 09-cab. Phylogenetic analyses of the gGAPDH and SSU rRNA genes showed that the flagellate within Lu. (H.) caballeroi (08-cab3, 08-cab4, and 09-cab) was located in the clade comprising T. avium isolated from chaffinch, lesser-spotted eagle (APO1), kestrel (FT2), fringuello (LSHTM 144B), and the black fly Eusimulium securiforme (SIM3) (Fig. 2A, B). 08-cab3, 08-cab4, and 09-cab were separated from other avian trypanosomes, T. corvi and isolates from mosquito (T. sp. CUL1) and hippoboscid fly (T. sp. OA6), and anuran Trypanosoma species isolated from sand flies in the Brazilian Amazon (T. sp. 101, 103, 120, 887, 888 and 1155) and in a desert area of Pakistan (T. sp. SKF32) (Fig. 2A, B). These results indicated that the flagellates within Lu. (H.) caballeroi were T. avium. Isolation of the flagellates was attempted in 09-per and 09-cab, and both parasites were successfully isolated in Grace's medium cocultured with Sf9 insect cells. The parasites in the culture were confirmed to correspond to the parasites detected within sand flies by the cyt b, gGAPDH, and SSU rRNA gene analyses.
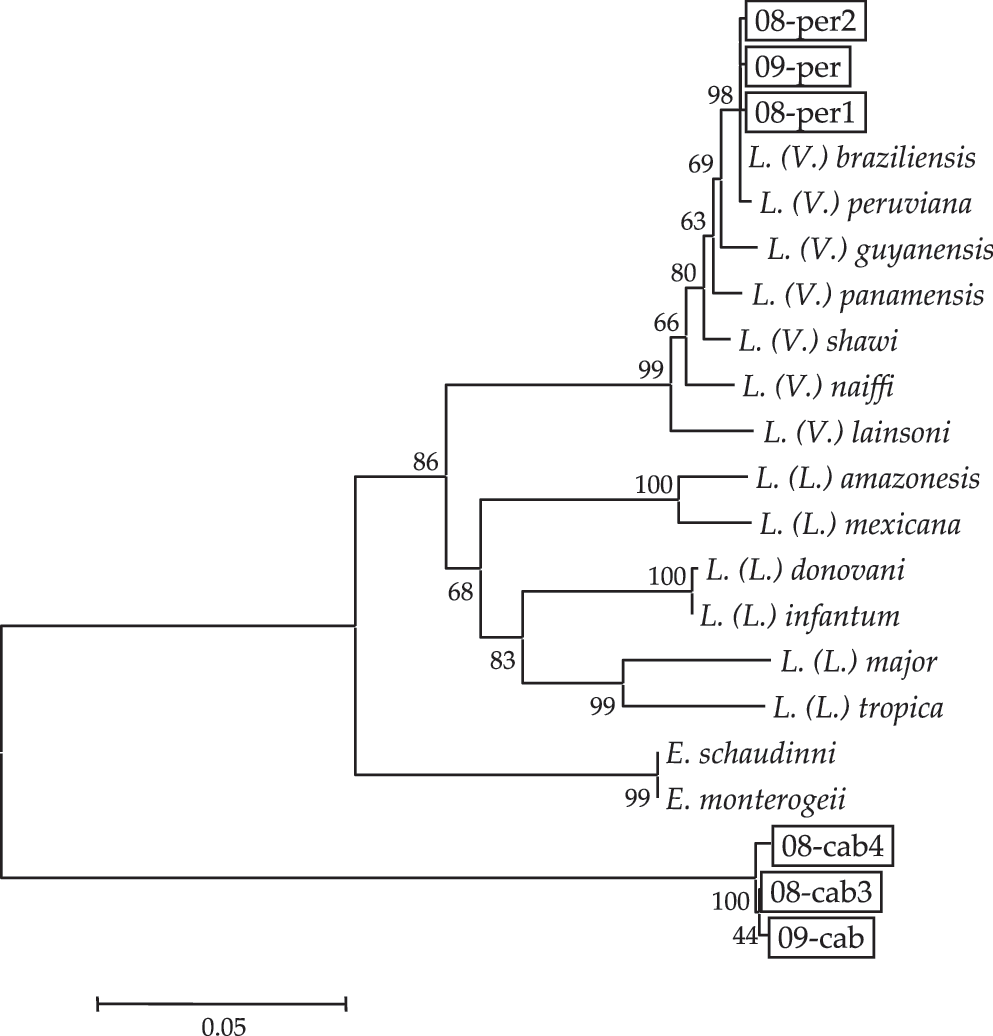
Phylogenetic tree of cyt b gene sequences among species. The cyt b genes were amplified from flagellates within Lutzomyia (Helcocyrtomyia) peruensis (08-per1, 08-per2, and 09-per) and Lutzomyia (Helcocyrtomyia) caballeroi (08-cab3, 08-cab4, and 09-cab) and sequenced. Phylogenetic analyses of the cyt b gene sequences were performed by the neighbor-joining method together with sequences from 13 Leishmania and 2 Endotrypanum species. The scale bar represents 0.05% divergence. Bootstrap values are shown above or below branches. cyt b, cytochrome b.

Phylogenetic tree of gGAPDH
Sand flies were captured around a patient's house at Nambuque where dogs, guinea pigs, chickens and turkeys were raised and in a farm at Padregual where field rodents, raptors, and other birds were found. The natural hosts of Le. (V.) peruviana and T. avium identified within sand flies were examined; however, no flagellates were observed in the peripheral blood of chickens and turkeys, and no leishmanial DNA was detected in the peripheral blood of dogs and various organs including liver, spleen, and lymph nodes of guinea pigs and field rodents in the present study.
Discussion
In the present study, the natural infection of sand flies by Leishmania species was examined in the northern Andean areas of Peru where CL caused by Le. (V.) peruviana is endemic, to define the vector species responsible for the transmission of the disease in these regions. Lu. (H.) peruensis and Lu. (H.) caballeroi were identified as the dominant man-biting species, and natural infections by flagellates were detected in both. Le. (V.) peruviana was identified within Lu. (H.) peruensis but not Lu. (H.) caballeroi by cyt b and MPI gene analyses, indicating the vector species responsible for the transmission of Le. (V.) peruviana in these areas to be Lu. (H.) peruensis. The flagellate within Lu. (H.) caballeroi was identified as T. avium on the basis of gGAPDH and SSU rRNA gene analyses. This is the first report of the infection of man-biting sand flies by trypanosomes other than Leishmania in Andean areas. Although many sand flies are opportunistic feeders, Lu. (H.) peruensis and Lu. (H.) caballeroi are considered to be highly man-biting species because both were efficiently captured by protected human bait. Blood-meal sources other than humans of these sand flies are under investigation to determine the natural hosts of Le. (V.) peruviana and T. avium.
Areas where CL caused by Le. (V.) peruviana is endemic are widely prevalent throughout the Andean cordillera in Peru (Lucas et al. 1998, Davies et al. 2000, Arevalo et al. 2007, Kato et al. 2010c). The sand fly species responsible for the transmission of Le. (V.) peruviana in nature have been identified as Lu. (H.) peruensis in the Huayllacallan valley, southern Ancash, and in Chaute, Department of Lima (Perez et al. 1994, 2007, Kato et al. 2008), and Lu. (H.) ayacuchensis in the western Andean valleys of Ayacucho (Caceres et al. 2004), all of which are located in the central Peruvian Andes. In this study, the natural infection of sand flies by Leishmania was examined in endemic areas of La Libertad located in the northern Peruvian highlands to further define vector species responsible for the transmission of Andean leishmaniasis. Flagellates were detected in two dominant sand fly species, Lu. (H.) peruensis and Lu. (H.) caballeroi, and 3 of the 211 (1.4%) Lu. (H.) peruensis specimens were infected by Le. (V.) peruviana, the causative agent for CL in these areas. The infection rate was higher than in previous reports; 1 of 75 Lu. (H.) peruensis (1.3%) was positive for Le. (V.) peruviana (Perez et al. 2007), 1 of 462 Lu. (H.) peruensis (0.22%) was positive for Le. (V.) peruviana (Kato et al. 2008), and 5 of 1849 Lu. (H.) ayacuchensis (0.27%) were positive for Leishmania species, 4 of which were identified as Le. (V.) peruviana (Caceres et al. 2004); however, infection rates could be affected by several reasons including transmission endemicity, seasonality, and methods for sand fly collection and detection of the infection. Another important finding is the detection of a high rate of T. avium infection (2.6%) within a man-biting sand fly, Lu. (H.) caballeroi. More than 100 species of avian trypanosomes including T. avium have been described so far (Votýpka et al. 2002, 2004, Votýpka and Svobodova 2004), and a wide variety of blood-sucking arthropods (mites, hippoboscids, biting-midges, black flies, and mosquitoes) have been described as vectors (Bennett 1970, Baker 1976, Molyneux 1977). However, to our knowledge, no phlebotomine sand fly has been reported to transmit avian trypanosomes. T. avium has been isolated from a black fly attacking the nestlings of raptors, and the transmission of T. avium from black flies to canaries was indicated experimentally (Votýpka et al. 2002). Although trypanosomes were isolated from mosquitoes, hippoboscid flies, and biting-midges collected simultaneously in the same study, the parasites were not T. avium, suggesting that these insects cannot support the growth of T. avium (Votýpka et al. 2002). In the present study, numerous flagellates identified as T. avium were observed in the hindgut of Lu. (H.) caballeroi, strongly suggesting that the sand flies support the growth of and transmit the parasites. Several sand fly species were reported to transmit Trypanosoma species in tropical and subtropical areas, and all the parasites, except for an opossum trypanosome, T. freitasi, transmitted by a mammal-feeding Lutzomyia (Psychodopygus) claustrei in the Brazilian Amazon (Naiff et al. 1989), were anuran trypanosomes (Anderson and Ayala 1968, Ayala 1971, Ayala and McKay 1971, Ferreira et al. 2008, Lemos et al. 2008, Viola et al. 2008, Kato et al. 2010b). Other than Trypanosoma species, nonpathogenic trypanosomes such as Endotrypanum, Crithidia, Blastocrithidia, Herpetomonas, and Phytomonas have been detected within man-biting sand flies in tropical and subtropical areas by using molecular biological methods (Kato et al. 2007, Rocha et al. 2010). Additionally, uncharacterized flagellates were found within field mice-feeding Lu. (H.) noguchii in an Andean area (Caceres et al. 2004). PCR-based molecular biological method is now a powerful tool for vector research on leishmaniasis because of its sensitivity and simplicity (Perez et al. 1994, 2007, Caceres et al. 2004, Kato et al. 2007, 2008, 2010a, Rocha et al. 2010). However, only the detection of the parasite DNA within sand flies is not sufficient to show the infection of Leishmania species, and further genetic analysis such as sequencing of the amplified DNA fragments would be essential for the vector research on leishmaniasis because the genetic characteristics of Leishmania are similar to those of other trypanosomes. In addition, extensive research on vector ecology, such as feeding preference, would be required to ensure the vector species, because the presence of leishmanial DNA within a sand fly is not a sufficient evidence to prove the transmission of Leishmania.
In conclusion, the natural infection of sand flies by Leishmania was microscopically examined in the northern Peruvian Andes where CL caused by Le. (V.) peruviana is endemic, and flagellates detected within Lu. (H.) peruensis were identified as Le. (V.) peruviana by molecular biological methods, strongly suggesting that the vector species responsible for the transmission of leishmaniasis is Lu. (H.) peruensis in these areas. Further, for the first time, an avian trypanosome parasite, T. avium, was found in a highly man-biting sand fly, Lu. (H.) caballeroi. The presence of trypanosomes other than Leishmania shows that care is needed when identifying parasitic organisms within vector insects in areas where leishmaniasis is endemic, although simple and quick techniques based on molecular biological methods are being rapidly developed.
Footnotes
Acknowledgments
The authors thank Flavio-Valeriano Zambrano C. (Servicio Nacional de Erradicacion de la Malaria, Guayaquil, Ecuador) and Roberto Sud A. (Ministerio de Salud Publica y Asistencia Social, Guayaquil, Ecuador) for their technical assistance throughout the study. This study was financially supported by the Ministry of Education, Culture and Sports, Science, and Technology (MEXT) of Japan (grant nos. 14256004, 18256004, and 18780230).
Disclosure Statement
No competing financial interests exist.