
Abstract
The distributions of the tick vector, Ixodes scapularis, and of the etiologic agent of Lyme disease, Borrelia burgdorferi (Bb), have continued expanding in Illinois over the past 20 years, but the extent of their spread is not well known. The role of multiple habitats in the establishment and maintenance of I. scapularis and Bb at local scales is not well understood, and the use of integrated approaches to evaluate local scale dynamics is rare. We evaluated habitat diversity and temporal changes of I. scapularis occurrence and Bb infection within a natural area in Piatt County, Illinois, where I. scapularis were first detected in 2002. Small mammals were trapped and attached ticks were collected in young forest, prairie, mature forest, and flood plain sites from 2005 to 2009. Small mammal abundance, and the prevalence (% mammals infested), mean intensity (I. scapularis per infested mammal), and relative density (I. scapularis per mammal trapped) of I. scapularis were computed for each habitat type and compared. Immature I. scapularis were tested for Bb infection using polymerase chain reaction techniques. Out of 2446 trapped small mammals, 388 were infested with I. scapularis. The prairie had the highest diversity of small mammal hosts. Prevalence, mean intensity, and relative density of I. scapularis and prevalence of Bb infection were highest for the prairie and young forest sites; in the former, all infection was associated with the prairie vole, Microtus ochrogaster. The minimum Bb infection prevalence of on-host I. scapularis collected in the natural area was 14% (n=56). Unlike previous studies solely focused on forested areas and Peromyscus leucopus, our study is the first to provide evidence of I. scapularis collected from prairie habitat and other reservoir hosts, particularly M. ochrogaster.
Introduction
In the Midwest, populations of both the vector tick and spirochete were once limited to southeast Minnesota and northwest Wisconsin (Davis et al. 1984, Callister et al. 1988, 1991); however, the focal endemic area has expanded farther into Illinois (Bouseman et al. 1990, Cortinas and Kitron 2006) and the surrounding states (Pinger et al. 1996, Walker et al. 1998, Lingren et al. 2005, Hamer et al. 2007) in the past 20 years. In Illinois, Lyme disease has mainly been identified in the northern third of the state (Kitron et al. 1991, Nelson et al. 1991, Jobe et al. 2006, 2007). There were 108 reported human cases in 2008 compared to only 35 cases in 2000 (IDPH 2010).
The distribution and occurrence of I. scapularis is typically associated with environmental factors such as tick habitat suitability (Kitron et al. 1991, Guerra et al. 2002) and small mammal host abundance (Anderson et al. 1983, Brownstein et al. 2003). Suitable habitat for I. scapularis has been characterized by oak-dominated forested areas with sufficient leaf litter and moist, sandy soils (Kitron et al. 1992, Schulze et al. 1998). Favorable vegetation, soil, topography, and climate are necessary for successful questing, molting, diapause, and oviposition of I. scapularis (Guerra et al. 2002). Although the ecological and spatial determinants of variation in I. scapularis distribution and habitat use are not well understood at the local-scale level (Goodwin et al. 2001), fragmentation of habitat into small forest patches has been associated with decreased vertebrate species diversity and increased densities of P. leucopus (Nupp and Swihart 1998, Rosenblatt et al. 1999). In some areas, the net effect of decreasing patch size is an increasing fraction of I. scapularis feeding on a Bb-competent reservoir, thus increasing the risk for high nymphal infection prevalence (Ostfeld and Keesing 2000, Allan et al. 2003).
Illinois, once dominated by tall-grass prairie, is now characterized by a highly fragmented landscape where forest patches are surrounded by urban areas, remnant prairie patches, and agricultural lands (Rosenblatt et al. 1999). The spatial variation and dynamism of the Lyme disease system highlight the need for a more comprehensive approach to study I. scapularis and Bb habitat suitability in small natural areas (Killilea et al. 2008). Both decreased biodiversity and increased habitat fragmentation are occurring throughout Illinois with potential implications for shaping Lyme disease risk.
In this study, we evaluated habitat diversity and temporal changes of I. scapularis occurrence and the maintenance of Bb among small mammal hosts within a natural area in east-central Illinois. The objectives of this study were (1) to determine I. scapularis occurrence and Bb infection across four habitat types within the natural area, and (2) to evaluate differences in I. scapularis occurrence and Bb infection over a 5-year period.
Materials and Methods
Study area
The study was conducted between 2005 and 2009 in Robert Allerton Park (RAP), a 614 ha natural area owned by the University of Illinois located 6.4 km southwest of Monticello, in Piatt County, Illinois. Since initial detection of I. scapularis in RAP in 2002, this study represents the only I. scapularis surveillance effort conducted in Piatt County to date. The park is bisected by the Sangamon River; the south half is a natural area consisting of a river corridor, a flood plain, upland and bottomland forests, meadows, and prairie surrounded by an intensively farmed agricultural landscape (Fig. 1). Four sites representative of the habitats found within the park were selected. Site 1 is an upland young successional forest comprised of oak-hickory (Quercus-Carya) stands and heavy undergrowth (Wang et al. 2008). Site 2 is a restored tall-grass prairie mainly comprised of big bluestem and Indiangrass (Andropogon gerardii, Sorghastrum nutans). Site 3 is a mature successional forest comprised of old-growth oak-hickory (Quercus-Carya) stands. Site 4 is a floodplain of the Sangamon River dominated by silver maple and green ash (Acer saccharinum, Fraxinus pennsylvanica) (Wang et al. 2008).
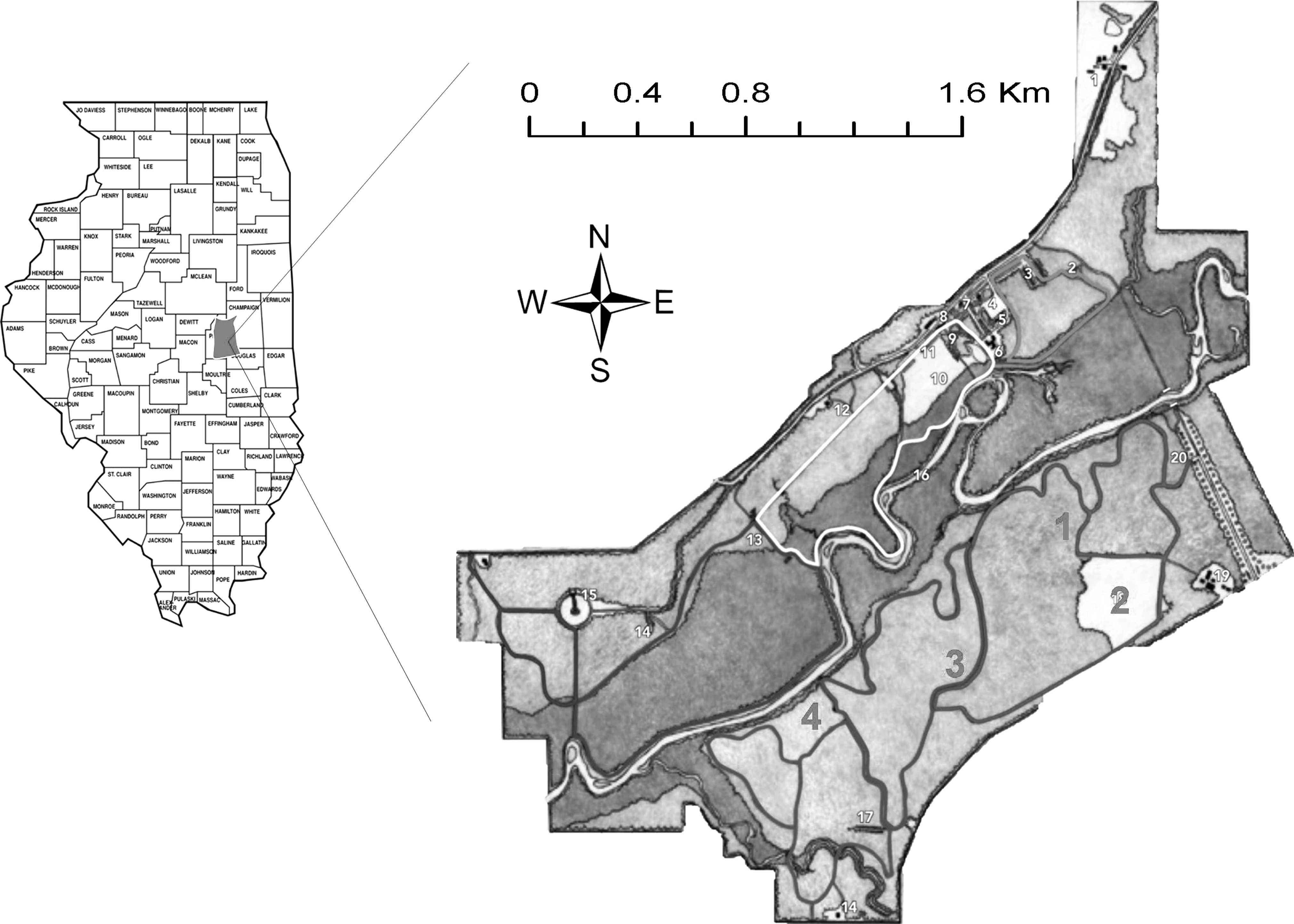
Study site in east-central Illinois. Robert Allerton Park (RAP) is located 6.4 km southwest of Monticello, in Piatt County, Illinois. Site 1=Young Forest, Site 2=Prairie, Site 3=Mature Forest, Site 4=Flood Plain.
Sampling frame
A 90×90 m grid consisting of 100 trapping stations set 10 m apart was established within each site similar to the methods described by Kitron et al. (1991). Two 5.08×6.35×16.51 cm3 Sherman live traps (H.B. Sherman Traps, Tallahassee, FL) baited with sunflower seeds were placed 1 m apart at each trapping station for a total of 200 traps per night at each site. Traps were set in the late afternoon and retrieved the following morning. Trapping took place approximately one night every 2 weeks from June through October 2005–2009 with the exception of two trap nights in May 2007 and one in November 2008. Each site was trapped between 3 and 11 nights per year (mean=6.3, SD=2.56), and between 30 and 33 nights over the study period (mean=31.5, SD=1.29).
Small mammal processing
Captured mammals were restrained, identified, ear-tagged (National Band and Tag, Newport, KY), and examined for ticks. Ticks were removed using tweezers and placed in vials containing 70% ethanol for later identification and lab analysis. Recaptured mammals were similarly processed, and their ear tag numbers were recorded. After examination, the small mammals were released at the trapping site.
Lab analysis
Identification of the ticks to life stage and species was performed under a dissecting microscope using an identification key (Sonenshine 1979). Multiple I. scapularis larvae from a single host were pooled and tested as one unit, and nymphs were tested individually. Total DNA from the larval pools and nymphs was extracted using the DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA) following a modified version of the Qiagen supplementary protocol: Purification of Total DNA from Ticks for Detection of Borrelia DNA.
The DNA samples were tested for Bb at Michigan State University using either (1) a nested polymerase chain reaction (PCR) for the 16S–23S rRNA intergenic spacer region of Borrelia spp. (Bunikis et al. 2004) followed by visualization with gel electrophoresis or (2) a quantitative PCR of a region of the 16S rRNA of Bb (Tsao et al. 2004). The intergenic spacer region primers used for PCR will amplify other Borrelia species. However, the only other two Midwest species detected in the lab at MSU are unlikely to be the cause of the positives. Borrelia miyamotoi makes a visibly smaller band on the gel, and Borrelia andersoni is bird-associated and has only been isolated from ticks removed from birds. A subset of the DNA samples were sequenced to confirm identification of Bb. The nested PCR was utilized for the first 2 years because these methods were already in place at MSU for use in other studies. The remaining 3 years of samples were tested using quantitative PCR, which is faster and more cost-effective. Preliminary experiments showed that both tests were comparable in sensitivity and were able to detect positive samples containing a minimum 100–10–1 organisms.
Statistical analysis
The number of captured small mammals and collected I. scapularis were calculated by habitat and by year. We used three measures to evaluate temporal changes of I. scapularis occurrence in each habitat: prevalence of I. scapularis infestation (proportion of mammals that were infested), mean intensity of infestation (number of I. scapularis per infested mammal), and I. scapularis relative density (number of I. scapularis per mammal). Prevalence of Bb infection in collected I. scapularis was quantified by the proportion of positive larval pools and nymphs from infested mammals. Differences in number of small mammals, number of I. scapularis, mean intensity of infestation, and I. scapularis relative density across habitats and years were compared using negative binomial regression. Prevalence of I. scapularis infestation and Bb infection were evaluated using logistic regression, and odds ratios (OR) were used to quantify the difference. Statistical analyses were performed using SAS software (Version 9.1.3 [2002–2004]; Statistical Analysis System Institute Inc., Cary, NC). A p≤0.05 was considered significant in this study.
Results
Small mammal abundance and species richness
Overall trapping effort amounted to 25,200 trap nights from 2005 through 2009. A total of 2446 captured small mammals (including recaptures) were examined for ticks. The white-footed mouse (P. leucopus) comprised the vast majority (96%) of individuals captured in all habitats. Other species processed include the meadow jumping mouse (Zapus hudsonius) in the young forest, flood plain, and prairie, and the prairie vole (Microtus ochrogaster) and the Western harvest mouse (Reithrodontomys megalotis) found only in the prairie. The abundance of small mammals captured was significantly different (p<0.001) across the four habitats. The abundance was highest for the flood plain (14.07 captures per 100 trap nights) followed by the mature forest (11.92 captures per 100 trap nights) and young forest (10.97 captures per 100 trap nights), then the prairie (2.17 captures per 100 trap nights). Despite the prairie having the lowest abundance of mammals captured, it had the highest species richness (Table 1).
Ratio of mean number of mammals captured per trap night, adjusting for year and month. Young Forest served as the reference group.
Ratio of mean number of Ixodes scapularis collected per trap night, adjusting for year and month. Young Forest served as the reference group.
I. scapularis occurrence
A total of 1009 immature I. scapularis ticks (977 larvae and 32 nymphs) were collected from the small mammals (Table 1). The number of I. scapularis collected was different (p<0.001) across the four habitats. The young forest had the highest number of I. scapularis collected (n=676), followed by the prairie (n=176), the mature forest (n=127), and the flood plain (n=30).
Prevalence of I. scapularis infestation did not vary between the young forest and the prairie (OR=1.0; 95% CI=0.7, 1.5) (Table 2). However, prevalence of I. scapularis infestation was higher in the young forest than in the mature forest (OR=0.2; 95% CI=0.16, 0.28) and the flood plain (OR=0.03; 95% CI=0.02, 0.05) (Table 2). Intensity of infestation showed the same pattern with the greatest number of I. scapularis per infested mammal found in the prairie and the young forest. These two habitats had, on average, twice as high an intensity of infestation as both the mature forest and the flood plain (Table 2). Based on the outcome of the previous measures, the density of I. scapularis was highest for the prairie and the young forest, followed by the mature forest, and finally the flood plain (Table 2).
Prevalence=infested mammals/trapped mammals, n is number of trapped mammals.
Odds Ratio is value per mammal, Ratio is value per trap night, both adjusting for year and month. Young Forest served as the reference group.
Mean Intensity=I. scapularis/infested mammals. I is number of infested mammals.
Relative Density=I. scapularis/trapped mammals.
Value significantly lower than young forest and prairie (p≤0.001).
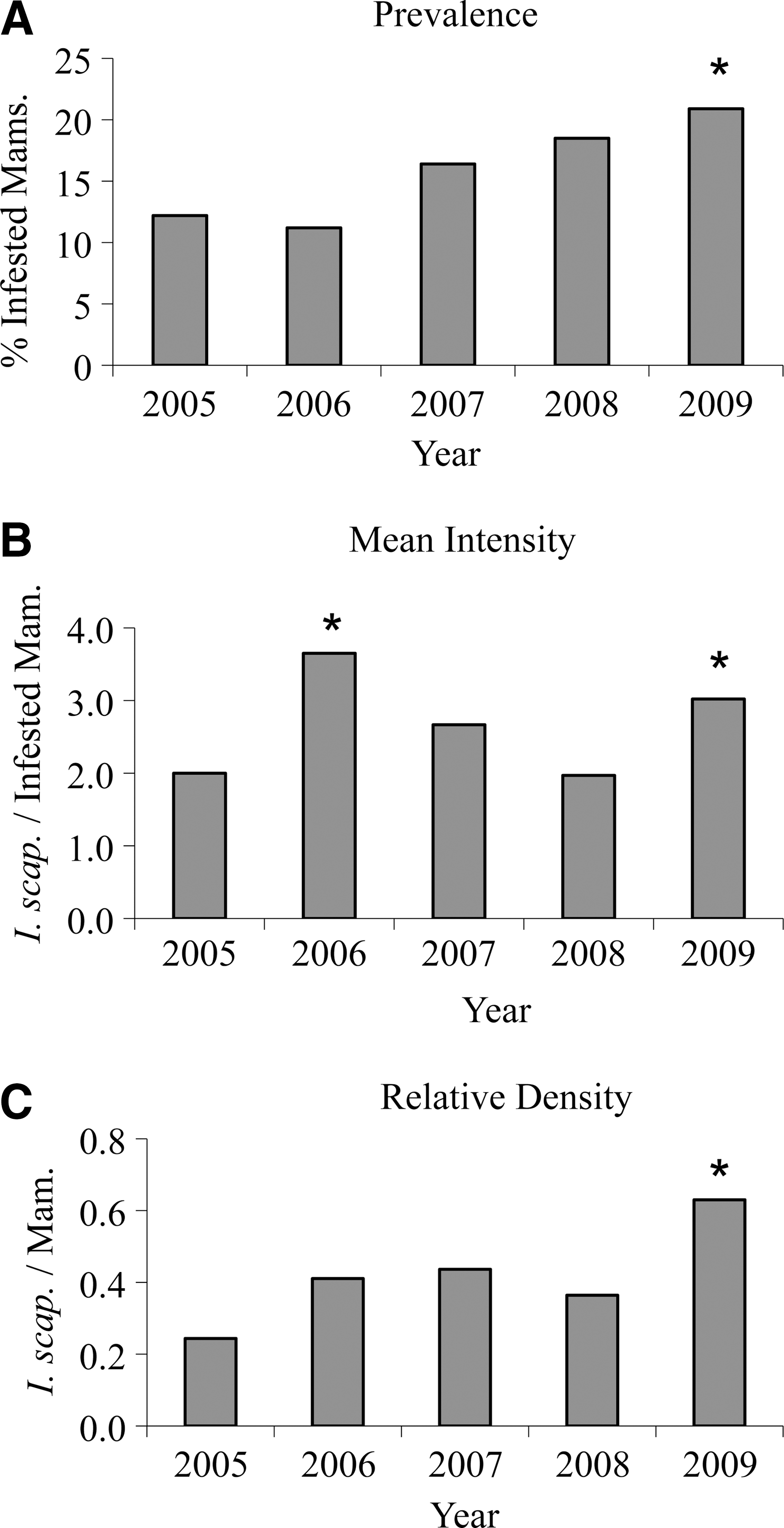
There was an increasing trend (p<0.001) in the prevalence of I. scapularis infestation over the study period (Fig. 2). The most recent year, 2009, had the highest prevalence (21%) of infestation and relative density (0.6 I. scapularis per mammal), and the second highest mean intensity (3.0 I. scapularis per infested mammal) (Fig. 2).
Bb infection
In total, 356 I. scapularis larval pools and 32 nymphs were tested for Bb by PCR (n=388), of which 53 larval pools and 3 nymphs were positive (n=56), for an overall infection prevalence of 14% for collected I. scapularis (95% CI=11%, 18%) (Table 3). The 3 positive nymphs were removed from captured small mammals in the young forest. The highest proportion of positive I. scapularis was found in the prairie (27%; 95% CI=16%, 41%). Of additional interest in this study, all positive I. scapularis from the prairie were larval pools collected from the prairie vole, M. ochrogaster. The odds of I. scapularis testing positive in the prairie were twice as high as the young forest (OR=2.2; 95% CI=1.0, 4.5) (Table 3). The small numbers of I. scapularis testing positive for Bb in the mature forest (n=5) and the flood plain (n=1) made it difficult to draw conclusions from these two sites (Table 3). The prevalence of Bb infection within RAP did not significantly vary over the study period (p=0.992), despite a lack of detectable infection in 2006 (Fig. 3), and significant variation in abundance of I. scapularis across years (p<0.001).
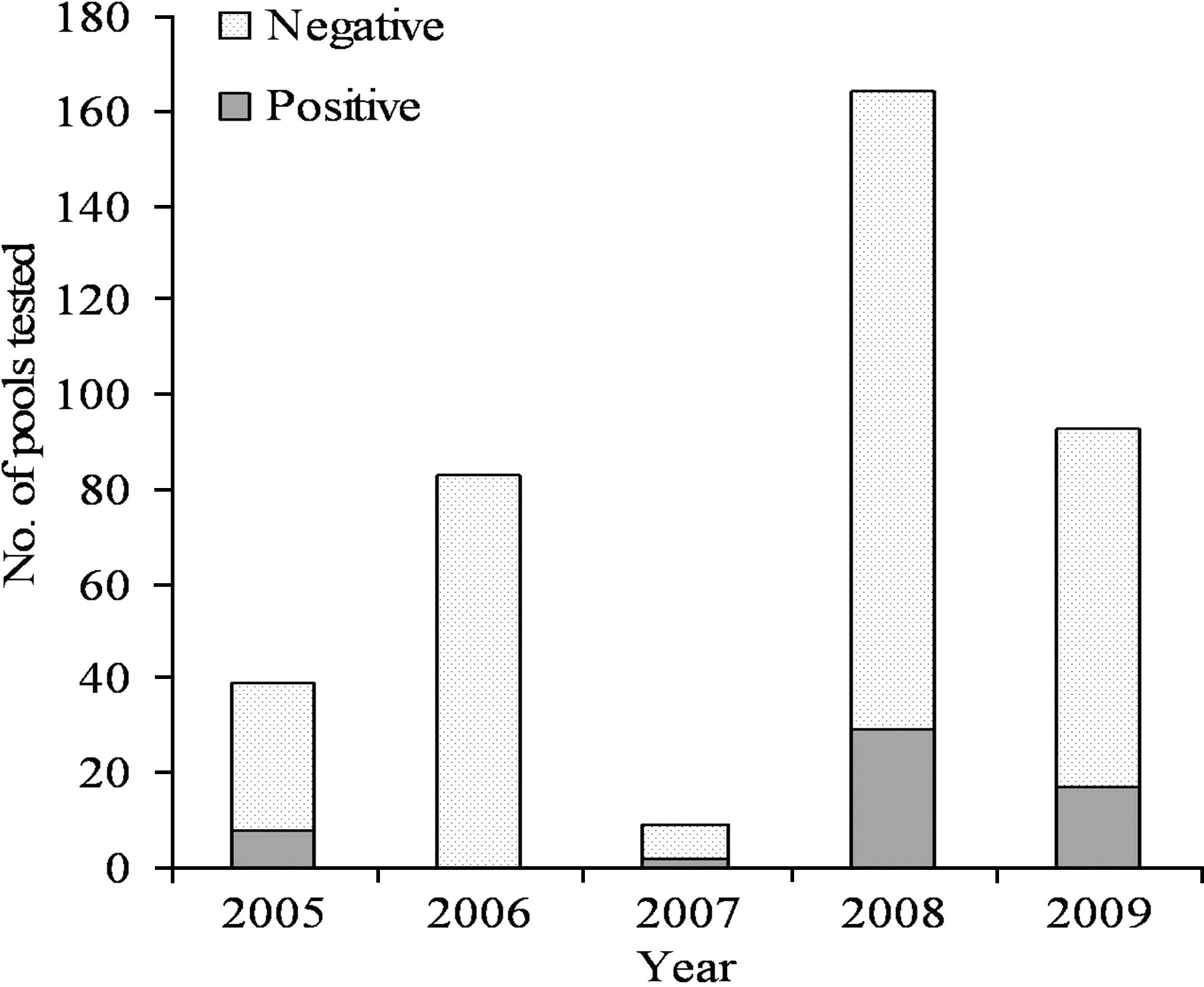
I. scapularis larval pools and nymphs tested for Borrelia burgdorferi in RAP, Piatt County, Illinois, 2005–2009. % positive I. scapularis did not vary significantly (p=0.992) across years.
Only three nymphs from the young forest tested positive for Bb.
Odds ratio of B. burgdorferi infection prevalence per infested mammal, adjusting for year and month. Young Forest served as the the reference group; ×, small sample size of Mature Forest (n=5) and Flood Plain (n=l) did not allow precise estimates.
Discussion
In the highly fragmented region of east-central Illinois, the Sangamon River is a vital riparian corridor among small forest and remnant prairie patches surrounded by agricultural fields and urban areas (Rosenblatt et al. 1999). This landscape produces a complex but relatively small network of habitat diversity within RAP with unknown effects on the dynamics of I. scapularis occurrence and Bb infection among small mammal hosts. The significant increasing trend (2005 to 2009) in prevalence of I. scapularis infestation in RAP in this study, compared to the initial reports in 2002 of 9 adult I. scapularis ticks from a deer-harvest county-surveillance effort, support the hypothesis that I. scapularis distribution continues to expand in Illinois (Cortinas and Kitron 2006, IDPH 2009). All three measures of I. scapularis occurrence were significantly higher in 2009, the final year of this study, resulting in an upward trend of infested hosts with higher tick burdens. If this pattern continues in coming years, an increase in Bb infection of I. scapularis in RAP is a conceivable possibility.
The data of this study indicate differences in I. scapularis prevalence, mean intensity, and relative density among habitats, with greater measures of I. scapularis occurrence in the prairie and young forest compared to both the mature forest and the flood plain. The prevalence of I. scapularis infestation among trapped small mammals ranged from 40% in the prairie to 33% in the young forest, 11% in the mature forest, and 2% in the flood plain. Although these results are comparable to small mammal studies in forest patches in northwestern Illinois (Kitron et al. 1991, Mannelli et al. 1994, Slajchert et al. 1997), this study is the first to report a prairie habitat supporting the highest prevalence of I. scapularis infestation compared to the forested sites within the same natural area. In addition, we are the first to report evidence that M. ochrogaster may potentially serve as a competent reservoir host for Bb. Further research is needed to verify the suitability of prairie habitat for I. scapularis population dynamics and of M. ochrogaster for Bb maintenance and transmission. If multiple habitat types populated by competent reservoir host species are suitable for supporting I. scapularis vectors, Lyme disease transmission could increase in RAP and in the east-central region of the state.
This study indicates that Bb is present in the park. This result is of pivotal importance to evaluate the risk of Lyme disease transmission in an area where the mechanisms of host–vector coexistence and the maintenance of Bb remain equivocal. The highest prevalence of Bb infection was found in on-host I. scapularis larval pools from the prairie (27%) followed by the young forest (15%), the mature forest (6%), and the flood plain (6%). Because of the small sample size of Bb-positive I. scapularis, more data are needed to increase precision of these point estimates.
Whereas I. scapularis is typically associated with sandy, oak, upland deciduous forest (Guerra et al. 2002), we found that the prairie had the highest occurrence of I. scapularis (although not statistically different from occurrence in the young forest), despite having the lowest abundance of small mammals. The prairie also had the highest observed species richness of small mammals, including Peromyscus spp. as well as M. ochrogaster, R. megalotis, and Z. hudsonius. These additional species may serve as substitute or additional hosts for I. scapularis where P. leucopus is less abundant (Mannelli et al. 1993, Markowski et al. 1997).
Although the meadow vole (Microtus pennsylvanicus) has been reported as a competent reservoir for Bb (Anderson 1988, Markowski et al. 1998), no reports to date describe M. ochrogaster as a potential reservoir host for Bb. Mather et al. (1989) reported that the prairie does not qualify as suitable habitat for I. scapularis populations, so the potential for small mammal reservoir hosts of Bb in this habitat (specifically M. ochrogaster in this study) may have been overlooked. All I. scapularis testing positive for Bb in the prairie were larvae collected from M. ochrogaster, which implies the larvae acquired infection from M. ochrogaster as their first blood meal. It must be noted that larval feeding can become interrupted and larvae may seek out an additional blood meal as a result. Nymphs could also acquire infection during a previous blood meal, but none were found on M. ochrogaster. Based on evidence from this study and the fact that M. ochrogaster and M. pennsylvanicus are closely related, it is plausible for M. ochrogaster to serve as a comptent zoonotic reservoir for Bb.
Previous studies suggest that decreased biodiversity and increased habitat fragmentation appear to have significant roles in the overpopulation of P. leucopus (Rosenblatt et al. 1999), the establishment of I. scapularis (Ginsberg 1994), and the risk of Lyme disease (Van Buskirk and Ostfeld 1995, LoGiudice et al. 2003). Conclusions from prior studies indicate that an increase in species diversity within host communities leads to a “dilution effect” of immature I. scapularis acquiring Bb infection because of more ticks feeding on inefficient reservoir hosts (Van Buskirk and Ostfeld 1995, Ostfeld and Keesing 2000). In other words, increased biodiversity of small mammals leads to I. scapularis feeding on a wider variety of hosts that have low Bb reservoir competency thus decreasing the chance of the feeding tick to become infected and reducing risk of human exposure to Lyme disease. The result is less infected I. scapularis in habitats with greater biodiversity. In contrast, our study shows an increase in I. scapularis occurrence and Bb infection in the prairie, where small mammal species richness is greatest. The presence of M. ochrogaster in the prairie may serve as a competent reservoir that is not found within other habitat types. In this manner, the vole may amplify the abundance of infected I. scapularis, as Ogden and Tsao (2009) alternatively suggest.
The observation made by Brown and Burgess (2001) that habitat heterogeneity leads to increased host diversity and increased rodent density may explain why the occurrence of I. scapularis is highest in the prairie as well as the young forest. Given the relatively small size of RAP (614 ha), I. scapularis occurrence and Bb prevalence may be dynamic, or continue to shift and change within and among the habitats of the park. The diverse habitats of this natural area are clustered within a close range and bordered by agriculture fields and urban areas (Rosenblatt et al. 1999). Such habitat composition favors a high degree of connectivity, enabling vertebrate host movements between suitable habitat patches, which, based on literature, is an important factor correlated with I. scapularis distribution and abundance (Estrada-Peña 2003). For example, this landscape could facilitate deer movement between habitats, enabling them to utilize the riparian corridor of the Sangamon River near the young forest as a dispersal route, the prairie as a bedding area, and the agriculture fields as a food source. Such deer habitat use could explain the high occurrence of I. scapularis in the prairie and young forest. As habitat patches become smaller in size due to changes in land use toward agriculture and urbanization, I. scapularis may increase in abundance and density in remaining patches (Ginsberg 1994).
We identified the occurrence of I. scapularis within habitats other than forested areas. These findings are supported by a number of exceptions to generalized tick–habitat associations (Piesman and Gray 1994), and the concept that landscape context can influence the abundance of I. scapularis (Madhav et al. 2004, Estrada-Peña 2009). Given the caution associated with surveying only one study site, our findings reveal a need to further investigate potential suitability of habitats other than forested areas and reservoir hosts, particularly M. ochrogaster, for I. scapularis population establishment and Bb transmission at a local scale in a dynamic fragmented landscape.
Footnotes
Acknowledgments
The authors acknowledge M. Buhnerkempe, J. Teisburg, L. Kammin, and S. Rao for their contributions to the initial stages of this study; N. Beccue, D. Becker, and Allerton Park Staff for access to study sites and facilities; and field and lab technicians and volunteers for data collection. The authors also thank Drs. L. Haramis and J. Nelson for providing valuable input to this study. Funding for this project was provided by the University of Illinois at Urbana-Champaign: Earth and Society Initiative, James Scholar Program, Department of Natural Resources and Environmental Sciences, Illinois Natural History Survey, and the Federal Aid in Wildlife Restoration Project W-146-R. This project was conducted under protocols approved by University of Illinois Biosafety and Institutional Animal Care and Use protocols.
Disclosure Statement
No competing financial interests exist.