
Abstract
The genus Orthopoxvirus includes variola (smallpox) virus and zoonotic cowpox virus (CPXV). All orthopoxviruses (OPV) are serologically cross-reactive and cross-protective, and after the cessation of smallpox vaccination, CPXV and other OPV infections represent an emerging threat to human health. In this respect CPXV, with its reservoir in asymptomatically infected wild rodents, is of special importance. In Europe, clinical cowpox has been diagnosed in both humans and animals. The main objective of this study was to elucidate the prevalence of OPV infections in wild rodents in different parts of Eurasia and to compare the performance of three real-time polymerase chain reaction (PCR) methods in detecting OPV DNA in wildlife samples. We investigated 962 wild rodents from Northern Europe (Finland), Central Europe (Germany), and Northern Asia (Siberia, Russia) for the presence of OPV antibodies. According to a CPXV antigen-based immunofluorescence assay, animals from 13 of the 17 locations (76%) showed antibodies. Mean seroprevalence was 33% in Finland (variation between locations 0%–69%), 32% in Germany (0%–43%), and 3.2% (0%–15%) in Siberia. We further screened tissue samples from 513 of the rodents for OPV DNA using up to three real-time PCRs. Three rodents from two German and one Finnish location were OPV DNA positive. The amplicons were 96% to 100% identical to available CPXV sequences. Further, we demonstrated OPV infections as far east as the Baikal region and occurring in hamster and two other rodent species, ones previously unnoticed as possible reservoir hosts. Based on serological and PCR findings, Eurasian wild rodents are frequently but nonpersistently infected with OPVs. Results from three real-time PCR methods were highly concordant. This study extends the geographic range and wildlife species diversity in which OPV (or CPXV) viruses are naturally circulating.
Introduction
Like humans, cats and possibly rats are incidental hosts for CPXV. Their skin lesions resemble those of humans (Bennett et al. 1989). Rats seem to die easily from cowpox (Marennikova 1979, Becker et al. 2009). Cats become infected by wild rodents, the reservoir of CPXV—that show no clinical signs (Bennett et al. 1997, Telfer et al. 2002).
OPV-specific antibodies are reported from wild rodents in England (Crouch et al. 1995); on the European mainland, Poland (Marennikova et al. 1984), Belgium (Boulanger et al. 1996), Norway (Tryland et al. 1998), Finland (Pelkonen et al. 2003), the Netherlands (Martina et al. 2006), and southern Germany (Essbauer et al. 2009); and in Central and Western Asia, Turkmenistan (Marennikova et al. 1978) and Georgia (Tsanava et al. 1989), but not in northern Asia. Apart from recent findings in zoos (Kurth et al. 2008), few serological findings have been confirmed by virus isolation or (conventional) polymerase chain reaction (PCR) (Essbauer et al. 2009). Several quantitative real-time PCR (qPCR) methods allow detection of OPVs (Nitsche et al. 2004, Olson et al. 2004, Scaramozzino et al. 2007, Putkuri et al. 2009), but reports on their use for wildlife samples or their comparison with each other are thus far unavailable.
To study how common are OPV infections throughout Eurasia, we screened wild rodents from Finland, Germany, and Siberia's Baikal region for OPV antibodies and searched for OPV DNA with qPCR followed by sequence analysis, and compared the performance of three qPCR methods.
Materials and Methods
Rodents
From several locations across Eurasia, we snap-trapped 962 wild rodents of 15 species, all without obvious clinical signs, according to national regulations (Fig. 1 and Tables 1 and 2). In total, 301 bank voles (Myodes glareolus) came from six locations in Finland, 224 rodents of six species from five locations in Germany (Ulrich et al. 2008), and 437 rodents of nine species from six locations in the Baikal region in the Republic of Buryatia, Siberia, and Russia (later “Buryatia”; Plyusnina et al. 2008) (Fig. 1). Snap-traps were checked daily, and carcasses were either frozen until preparation (Finland and Germany) or prepared the same day and stored in RNAlater Stabilization Reagent (QIAGEN), then frozen 2 to 10 days later (Buryatia). At necropsy, species and sex were recorded, and samples collected for PCR (lung: Finland, Buryatia; liver: Germany) and for serology. Blood samples were obtained from the thoracic cavity by either rinsing the cavity with phosphate-buffered saline (PBS; Germany, Ulrich et al. 2008), placing the heart in PBS (Finland; Sironen et al. 2002), or soaking blood into a filter paper, later dissolved out from the paper in PBS (Buryatia; Plyusnina et al. 2008). All samples were stored at −20°C until further use.
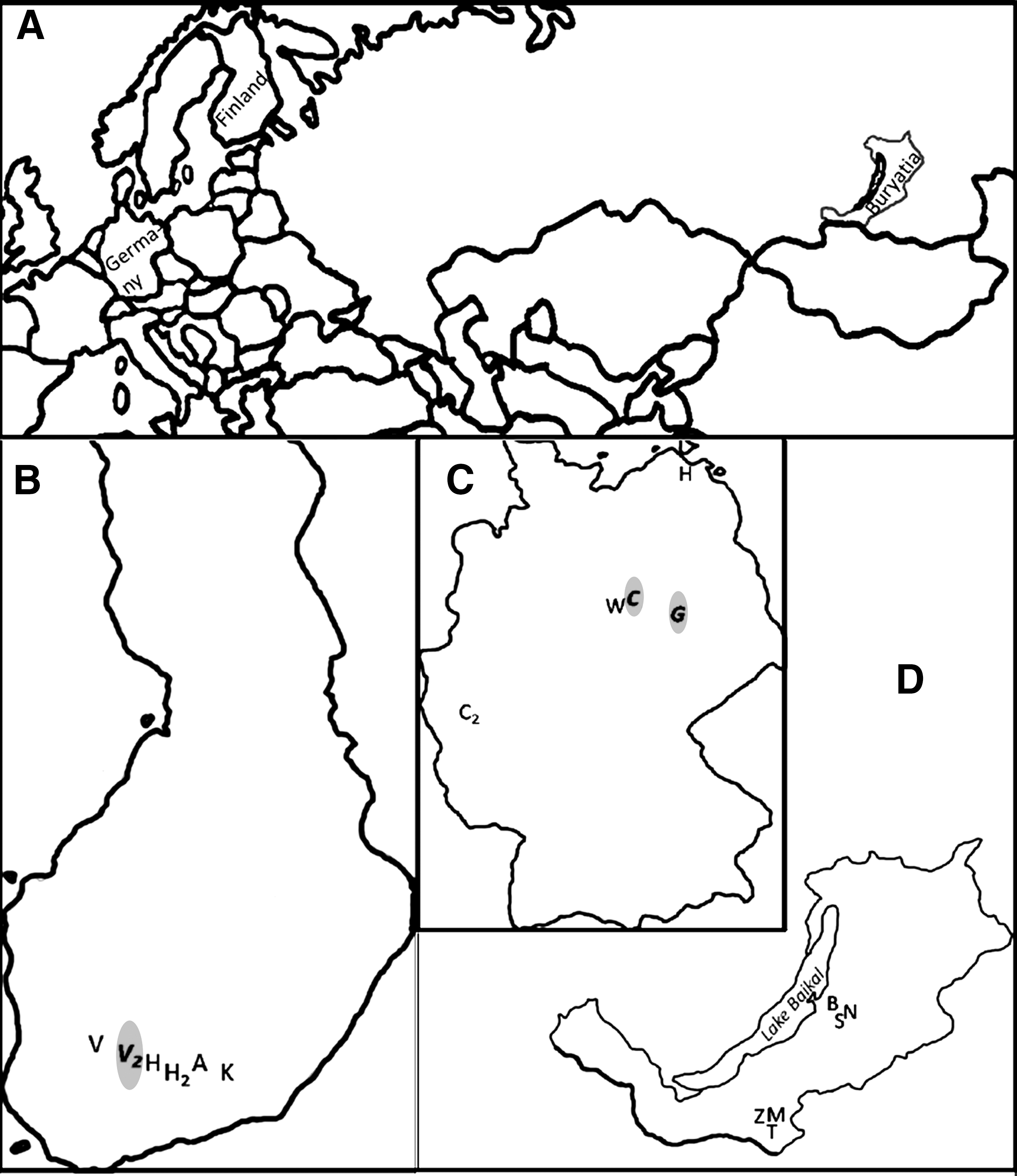
Rodent-trapping locations across Eurasia. Locations of orthopoxvirus DNA detection are highlighted in
IgG antibodies measured from blood samples by indirect immunofluorescence assay (IFA) with cowpox virus CPXV/FIN/T2000 as antigen (Pelkonen et al. 2003).
All the species names according to Wilson and Reeder 2005.
IgG antibodies measured from blood samples by IFA with cowpox virus CPXV/FIN/T2000 as antigen (Pelkonen et al. 2003).
All samples analyzed with a real-time PCR that amplified the partial orthopoxvirus hemagglutinin gene (Putkuri et al. 2009). German samples and PCR-positive Finnish samples were also tested with two other real-time PCRs, which targeted the 14 kDa protein gene (Olson et al. 2004 and Scaramozzino et al. 2007).
Confidence intervals for prevalence, Mid-P Exact.
Species names according to Wilson and Reeder 2005. M. glareolus, Myodes glareolus (bank vole); M. agrestis, Microtus agrestis (field vole); A. agrarius, Apodemus agrarius (striped field mouse); M. fortis, Microtus fortis (reed vole); M. rufocanus, Myodes rufocanus (gray red-backed vole); M. oeconomus, Microtus oeconomus (root/tundra vole); C. barabensis, Cricetulus barabensis (striped dwarf hamster).
Localizations as federal state of Germany, district, trapping site: MWP, Mecklenburg-Western Pomerania; NRW, North Rhine Westphalia; SA, Saxony-Anhalt.
PCR, polymerase chain reaction.
Serology
OPV-specific IgG antibodies were detected from the blood samples by indirect immunofluorescence assay (IFA), with CPXV/FIN/T2000 as antigen (Pelkonen et al. 2003, Kinnunen et al. 2007). Briefly, samples and subsequently mouse-Ig-specific fluorescein isothiocyanate-conjugated secondary antibodies (DAKO) were incubated on CPXV-infected as well as uninfected control cells at 37°C for 30 min and washed 3× with PBS after each incubation step. To estimate duration of antibody response, the avidity of OPV antibodies was measured by washing with PBS containing 6-M urea to detach low-avid IgG from the antigen, and the avidity index (AIX) calculated [AIX = (IFA titer with urea wash)/(IFA titer without urea wash) × 100%] (Hedman et al. 1991, Gavrilovskaya et al. 1993, reviewed in: Hedman et al. 1993). Samples with AIX ≥25% were interpreted to contain antibodies of high avidity, that is, strongly antigen-bound Ig-molecules, indicating adaptive immunity due to an OPV infection occurring weeks to years previously (Gavrilovskaya et al. 1993, Hedman et al. 1993, Pelkonen et al. 2003). The IFA and AIX measurements, originally established for human sera but also used for hantavirus-infected rodents (Gavrilovskaya et al. 1993, Pelkonen et al. 2003), were verified for CPXV-infected rodents by testing sera from two experimentally intranasally infected, sickened but recovered laboratory rats and a control rat. Antibody titers of 512 and 1024 and high avidity were measured in infected rats, whereas the noninfected rat's titer was <16.
Confidence intervals for prevalences were calculated by the Mid-P Exact test on the OpenEpi calculator (
Nucleic acid extraction and PCRs
A total of 513 rodents, including ones both seropositive and seronegative, were selected for PCR studies (Table 2). Total nucleic acids were extracted from homogenized tissue samples by commercial kits: the MagNA Pure total nucleic acid kit (Roche) on a MagNA Pure LC automaton for 4 mg of lung of the Finnish rodents, the NucleoSpin 96 Virus Core Kit (Macherey-Nagel) on a Hamilton automaton for 20 mg of liver of the German rodents, and the AllPrep DNA/RNA mini kit (QIAGEN) for 10 mg of lung of the Buryatian rodents. Nucleic acid extraction was repeated for PCR-positive and some PCR-negative samples with the High Pure PCR Template Preparation Kit (Roche).
The samples were analyzed with a qPCR targeting the partial OPV hemagglutinin (HA) gene with an analytical sensitivity (Se) of 1.3 to 13 genome copies (“PCR-1”; Putkuri et al. 2009). The 198 German samples and the 1 PCR-positive Finnish sample were also tested with two 14 kDa fusion protein gene-specific qPCRs: “PCR-2” (Se = 4–6 copies; Olson et al. 2004), and “PCR-3” (Se = 25 copies; Scaramozzino et al. 2007). In PCR-2 and −3, we controlled extraction and amplification using a heterologous internal control system, an in vitro transcribed HEX-labeled enhanced green fluorescent protein gene added to the reaction mixture (Hoffmann et al. 2006).
For all samples positive in qPCR, conventional OPV PCRs were used in attempted amplification of larger parts of the genome from several genes: whole A-type inclusion body (Meyer et al. 1994), partial thymidine kinase (Sandvik et al. 1998), and the whole (Damaso et al. 2000) as well as partial HA gene (artus Orthopox LC PCR kit; QIAGEN).
To prevent contamination, we separated the pre- and post-PCR stages of our work in different buildings. To monitor potential cross-contamination, we had already included 14 liver samples from specific pathogen-free mice as controls during the homogenization step and added respective negative controls to each qPCR run. A diluted plasmid containing 120 copies of partial VACV HA gene per μL (PCR-1; Putkuri et al. 2009) or 500 genome equivalents of modified VACV Ankara DNA (PCR-2 and -3; Timm et al. 2006) served as positive controls.
Sequence determination and analysis
The amplicons were purified with the QIAquick Gel Extraction kit (QIAGEN), and cycle sequenced in both directions with Big Dye Terminator reagents (Applied Biosystems) on an ABI 3130xl capillary device. The sequences were analyzed with the BioEdit program (
Results
OPV antibodies
Of the 962 wild rodents, 183 carried OPV antibodies (Table 2). Mean seroprevalence was 33% (95% CI 27%–38%; variation between locations 0%–69%) in Finland, 32% (26%–38%; 0%–43%) in Germany, and 3.2% (1.8%–5.2%; 0%–15%) in Buryatia. Antibodies appeared in 76% (13/17) of the locations and in seven rodent species. The avidity of antibodies was analyzed for 58 seropositive individuals with larger sample volume—of whom, 45 (78%) contained high-avidity antibodies (Table 3; some data not shown).
Trapping sites in Germany as federal state, district, and trapping site, SA; Saxony-Anhalt.
Total seroprevalence at that trapping site.
Existence of cowpox virus/orthopox virus-specific antibodies as measured by IFA.
Avidity interpreted as high if (IFA titer with urea wash)/(IFA titer without urea wash) ×100% ≥25%.
PCR-1: Hemagglutinin gene (Putkuri et al. 2009).
PCR-2: 14 kDa protein gene (Olson et al. 2004).
PCR-3: 14 kDa protein gene (Scaramozzino et al. 2007).
Ct value: Mean of the PCR cycle, in which sample fluorescence crossed background fluorescence.
NA, not available.
OPV DNA
OPV DNA was sought by qPCR from tissue samples of 513 rodents originating from locations with antibody-positive animals (Table 2). Three bank voles were positive in at least two of the PCR assays (rodents A, B, and C; Table 3) and one vole in all three (rodent B). Confirmation of the results by independent DNA extractions from these three positive rodents showed consistent PCR results. Ct values of the OPV DNA-positive rodent samples ranged from 33.2 to 39.4 depending on assay and tissue type (Table 3); a 2- to 3-cycle difference appeared between each individual's liver and lung. In addition, liver samples from two bank voles from one study site [W, Saxony-Anhalt (SA) (Fig. 1), in Germany], expressed positivity in PCR-3 only, and one additional animal from that site gave a borderline result (very low fluorescence) in PCR-1. Since we were unable to verify these results with the other PCR assays, we did not score these animals as positive. The added internal controls were clearly amplified in all the reaction mixtures, so no inhibition was observable in the tissues. Of the 198 rodents investigated by all three qPCR methods, results were concordant for 193 (97%).
Only one of the conventional PCRs (artus Orthopox LC kit, “second system” for HA gene; Qiagen) amplified OPV DNA from the three qPCR-positive samples.
OPV sequences
We retrieved short HA sequences from the three PCR-positive voles. Because their sequences differed from those of the positive controls, contamination could be excluded. A 179-bp sequence (accession number HQ127621) from Finnish rodent A matched 100% with Finnish CPXV strains T2000 and E1989 (Pelkonen et al. 2003) and showed one base-pair mismatch to the positive control VACV. From rodents B and C from two trapping sites in SA, Germany, 132-bp sequences (HM991174 and HM991175) retrieved for comparison were 100% identical with each other and with German CPXV strains HA16 (AY902268), HA07 (AY902256), 2000/20780 (FJ944572), 2007/26903 (FJ944563), and 2007/37472 (FJ944574), as well as with several MPXVs. Identity with pet-rat CPXV strains circulating from 2008 to 2009 in Germany and France (for instance, GQ260461) was <97%. Of the three CPXV DNA-positive rodents, two (B and C) were also antibody-positive (Table 3).
Discussion
Here we report a large survey of rodent OPV antibody prevalence and the first OPV DNA findings from Finland and Germany, including the first evidence from OPV infections in the Baikal region, Buryatia. Earlier, the easternmost CPXV findings were from Turkmenistan, 3000 km southwest of Buryatia (Marennikova et al. 1978). Other researchers have found OPV-specific antibodies in wild rodents elsewhere in Eurasia, but few have sought or detected viral DNA (Essbauer et al. 2009). We detected OPV DNA in one Finnish and two German bank voles. OPV DNA was amplified either from two tissues, or by separate qPCRs amplifying different gene regions, or both (Table 3). The PCR-positive individuals belonged to panels with relatively high seropositivity (>18%), thus verifying the presence of OPV infections in these populations.
Although the analytical sensitivities of the three qPCR methods are quite similar (1.3–13/4–6/25 copies), the discordant results for two rodents from Germany might be explainable by differences in sensitivity of these assays in practice, for instance, effects of host tissue DNA and of PCR conditions. These effects play an especially significant role when the template amount approaches the limit of detection (Scaramozzino et al. 2007). As these two animals originated from SA, where the clear PCR positivity of rodents B and C confirmed the existence of ongoing infections, it is possible that their low viral load was detected by PCR-3 but was below the detection limit of less sensitive PCR-1 and -2. PCR-2 detected all the verified positive and negative individuals, with few discordant results from the other two tests: PCR-1 gave unclear results for one positive and one negative rodent, and PCR-3 missed one verified positive but scored two other individuals positive. Additional investigations should evaluate any practical sensitivity differences between these methods, because positive samples were few, and nucleic acid qualities might have differed.
Overall, few individuals were PCR positive when compared to the parallel seroprevalence analysis. Earlier, 15% DNA and 14% antibody (by IFA) prevalences have been reported in small, wild Norwegian mammals (Sandvik et al. 1998, Tryland et al. 1998). In England, 3.2% to 8.2% of particularly targeted seroconverting rodents have been PCR positive (Chantrey et al. 1999). At least in England, infection dynamics of CPXV depend on rodent density and number of susceptible animals, and periods have existed with very small proportions of infectious rodents (Begon et al. 2009, 2009b). We may have trapped in such a suboptimal season. In addition, as compared to the long persistence of OPV antibodies (Kim et al. 2007), the period of detectable CPXV in wild rodents is very short: from 1 to 5 weeks postinfection in experimental settings (Marennikova 1979, Chantrey et al. 1999), so our low OPV DNA prevalence is unsurprising. Moreover, the high antibody avidity shows that in most cases the OPV infection was not acute.
Even the PCR-positive rodents were probably not sampled during maximal viral loads, since positive individuals were few, Ct values were high, most conventional (insensitive) PCRs were unsuccessful, and virus isolation in Vero cells and in 22 newborn mice failed (data not shown). CPXV/OPV isolation has proven difficult from rodents (Crouch et al. 1995, Bennett et al. 1997, Tryland et al. 1998), although repeatedly successful from feline and human patients (Campe et al. 2009, Johnson et al. 2009). Despite the low overall PCR prevalence, OPV DNA and therefore active infection were demonstrable in 25% (3/12) of the locations containing antibody-positive rodents.
The bank vole, long-tailed field mouse (Apodemus sylvaticus, wood mouse), and field vole (Microtus agrestis) have been considered major reservoir hosts maintaining and transmitting CPXV (Chantrey et al. 1999, Smith et al. 2009). CPXV or its DNA is also detectable in other wild rodent species: the root vole (Microtus oeconomus; Marennikova 1979), yellow suslik (Citellus vulvus), gray red-backed vole (Myodes rufocanus), northern red-backed vole (Myodes rutilus), brown rat (Rattus norvegicus; Essbauer et al. 2009), and gerbils (Rhombomys opimus and Meriones spp.; Marennikova et al. 1978, Tsanava et al. 1989). Further, many other rodent species, including the Norway lemming (Lemmus lemmus), yellow-necked field mouse (Apodemus flavicollis), common vole (Microtus arvalis), and Eurasian water vole (Arvicola amphibius), as well as insectivores, may become infected, as based on OPV antibody findings, but their role in viral epidemiology is unclear (Tryland et al. 1998, Essbauer et al. 2009, 2010). In addition to detecting OPV antibodies from reportedly antibody-positive species, the bank vole, field vole, root vole, and gray red-backed vole (Table 1), we identified three new possible host species: the striped field mouse (Apodemus agrarius), reed vole (Microtus fortis), and, interestingly, the striped dwarf hamster (Cricetulus barabensis), closely related to a popular pet hamster (Cricetulus griseus). Prevalence was highest in the striped field mouse, a new potential reservoir species, followed by the field vole and bank vole, well-known CPXV hosts.
Local OPV antibody prevalences in Eurasia have varied greatly, occasionally even reaching 100% (Burthe et al. 2006, Essbauer et al. 2009). Our observations of prevalences ranging from 0% to 69% parallel this pattern—findings probably reflecting well the real situation in our locations at trapping time, because we selected CPXV as an antigen and used IFA, the standard assay (Fenner 1994, Crouch et al. 1995, Essbauer et al. 2009), the performance of which we verified with control rat sera. Because all OPV antibodies cross-react, it is conceivable that OPVs other than CPXV cause some of the positive antibody findings. Other rodent-infecting OPVs include MPXV and taterapox virus in Africa, volepox virus in North America, and ectromelia virus (ECTV) in laboratory mice in Europe, China, Japan, and the United States (Fenner 2000, Damon 2007). ECTV can be experimentally transmitted to unspecified wild rodents (Groppel 1962), but not to the bank vole, which, in contrast, is susceptible to CPXV (Bennett et al. 1997). Evidence of natural ECTV infections in wild rodents is lacking (Fenner 1994), as is any indication that a single rodent species can host more than one OPV (Damon 2007). Another OPV species, VACV, originally derived from the human vaccination process, has caused zoonotic outbreaks in Indian buffaloes and Brazilian cattle, and may have a wild rodent reservoir in Brazil (Abrahão et al. 2009, Essbauer et al. 2010). VACV has also been suspected as infecting European wild rodents, but evidence is lacking (Boulanger et al. 1996).
The close relationship of OPVs is reflected in our novel sequences: the two short sequences from German rodents, in addition to CPXV, were identical also to MPXV, although differing from other OPV species. CPXV has, however, likely caused all our findings because MPXV, taterapox virus, volepox virus, and VACV are nonendemic in Europe (Damon 2007, Essbauer et al. 2010), and all characterized zoonotic OPV infections derived from wildlife in Europe have resulted from CPXV (Hansen et al. 2009, Essbauer et al. 2010). Exact proof should be ensured by thorough genetic studies of further OPV DNA sampling among wild rodents.
In this study, we retrieved no longer sequences, presumably due to low DNA/virus levels, despite trials with four conventional PCRs. Moreover, reliable phylogenetic analysis of these sequences was unfeasible because of their shortness. However, the Finnish sequence was, interestingly, identical to sequences reported earlier from Finland, suggesting that few closely related CPXV strains circulate there. In contrast, CPXV strains from Germany suggest a higher variation, because our sequences matched viruses isolated from a human and four cats in southern Germany but differed from other CPXV sequences (Kaysser et al. 2009) and from the rat strains (Campe et al. 2009).
In conclusion, OPV infections are distributed quite widely: In all these regions and even in 76% of the locations at least some rodents exhibited OPV-specific antibodies. Active infections were demonstrated by OPV DNA in rodents from 25% of the antibody-positive sites. Investigations of rodent samples by three OPV qPCR methods resulted in highly concordant results. In summary, this study has extended both geographic range and species diversity regarding the issue of OPV (or CPXV) circulating in wildlife.
Footnotes
Acknowledgments
We acknowledge Tytti Manni, Leena Kostamovaara, and Pirjo Sarjakivi for excellent technical assistance; Galina B. Muryeva, Zoya Yampilova, and Ilkka Alitalo for coordination of trapping in Buryatia; Stanislav Suhunov, Anne Balkema-Buschmann, Daniel Balkema, Wolfgang Wegener, Lutz Ohlmeyer, Dörte Kaufmann, Marc Mertens, Robert Klopfleisch, Christina Maresch, Thomas Büchner, and Daniel Windolph for rodent collection and necropsy; Anu Jääskeläinen and Antti Kinnunen for help with figure preparation; and Donata Kalthoff, Martin H. Groschup, and Martin Beer for continuous support.
This work was funded by the University of Helsinki and the Finnish Graduate School of Applied Bioscience and supported by University of Helsinki grant TA 715321 (project 2103039), EU grant GOCE-CT-2003–010284 EDEN, the Foundation of Veterinary Sciences, the Foundation for Support of Veterinary Research, and the Finnish Concordia Fund. The article is catalogued by the EDEN Steering Committee as EDEN0244 (
Disclosure Statement
No competing financial interests exist for any author.