
Abstract
Successful implementation of an integrated vector control program will rely on availability of accurate vector information in the specific location. However, such information can be limited in some countries. The aim of this study was to obtain baseline vector information from Pointe Noire on the Congo coast (Republic of the Congo). Field sampling was conducted during April 2009 in the village of Boutoto and its surrounds, close to the city of Pointe Noire. Anopheles gambiae sensu lato mosquitoes were collected resting indoors. Samples were analyzed for insecticide susceptibility, species identification, and Plasmodium sporozoite infection. Molecular and biochemical assays were conducted to characterize insecticide resistance mechanisms. The malaria vector A. gambiae S-form was the only mosquito species identified, and it had a high Plasmodium falciparum infection rate (9.6%). Multiple insecticide resistance was detected in this population with full susceptibility to only one insecticide class, the organophosphates. Dieldrin and DDT resistance was mainly attributed to target-site resistance (the Rdl and L1014F/L1014S kdr mutations respectively), whereas pyrethroid resistance was mainly attributed to P450 metabolic enzyme-mediated detoxification in addition to kdr. The role of various insecticide resistance mechanisms revealed a complex association between metabolic detoxification and reduced target-site sensitivity.
Introduction
Malaria control, particularly in Africa, has received major attention in the last decade. Funding opportunities have also been made available to combat malaria through cross-border initiatives such as the Lubombo Spatial Development Initiative (Sharp et al. 2007a) or by multicountry initiatives such as the President's Malaria Initiative (
The Republic of the Congo is located in Central/West Africa and is also referred to as Congo-Brazzaville, after its capital. The country is bordered by Gabon, Cameroon, the Central African Republic (CAR), the Democratic Republic of the Congo (DRC), and Angola. The Congo became independent in August 1960. Soon after independence the country experienced various uprisings and political instabilities such as those of 1963, 1968, 1977, 1979, and 1997–2003. These challenging circumstances nullified the possibility of large-scale and sustained malaria control. As a result, there is limited information available regarding malaria vector species composition and susceptibility to insecticides. Malaria is present throughout the year; however, malaria transmission peaks during and just after the rainy season. The rainy season differs within the country, and north of the equator the peak season will be from April through October, whereas October through to May for those areas that are south of the equator. More than 90% of malaria cases are caused by Plasmodium falciparum (
Here we report on a baseline malaria vector survey in the vicinity of Pointe Noire that was conducted to identify vector species, their P. falciparum infectivity rates, and susceptibility to insecticides (DDT, deltamethrin, bendiocarb, dieldrin, and malathion). This information has been used for the planned implementation of a vector control program in the study area. In addition to this, we also try to elucidate the resistance mechanisms in this population to aid in future resistance management in this study site. We looked at the presence of mainly two resistance mechanisms: target-site insensitivity as well as metabolic-mediated resistance.
Materials and Methods
Sample collection
Field sampling was conducted during April 2009 in the village of Boutoto and its surrounds (4°40′31S; 11°58′14E) near the Kouilou Potash mining project in Pointe Noire. Mosquitoes were collected indoors using manual aspirators. Adults were morphologically identified as members of the Anopheles gambiae complex using the keys of Gillies and Coetzee (1987). Wild-caught females were used for insecticide susceptibility tests in the field (Table 1), and a portion of live females (n = 110) were transported back to South Africa. All females transported back to SA and were placed in individual vials for oviposition. Eggs were transferred into larval rearing bowls, and progeny from each family (isofemale lines) were reared to F1 adults. The F1 progeny were used to establish a laboratory colony, named Congo A. gambiae S-form (COGS). Adults that emerged from F1 progeny were allowed to mate (These adults were called F0 of the COGS colony). Adults were provided with 10% sugar solution and three blood meals per week. Egg plates were provided to adults twice a week and eggs oviposited was reared through to the form the next generation (F1), etc.
n = total number of mosquitoes exposed or analyzed.
Total number of exposed mosquitoes, excluding controls.
WHO insecticide susceptibility assays
Standard WHO insecticide tests were used to assess the insecticide susceptibility of field-collected material and laboratory-reared samples (COGS) (WHO 1998). Samples of field-collected adults were exposed to 4% DDT, 0.05% deltamethrin (or 0.75% permethrin and 0.05% lambda-cyhalothrin for the laboratory reared strain, COGS), 0.1% bendiocarb, 4% dieldrin, and 5% malathion for 1 h each (n = 413). Adult mosquitoes from the laboratory-reared COGS were collected 1–3 days postemergence and exposed to insecticides as mentioned above. Each test consisted of ∼25 mosquitoes per exposure tube with one unexposed control tube. At least two replicates were performed for each exposure set. The number of mosquitoes knocked down (mosquitoes that either died in the first hour of exposure or those mosquitoes that are alive, but not able to fly) after 1 h exposure was recorded, and final mortality was noted 24 h postexposure and used to calculate the percentage of mortality. Mosquitoes had access to a 10% sugar solution during the 24 h holding period. Each exposure test included an untreated control that contained 25 mosquitoes exposed to paper lacking the relevant insecticide. There was 100% survival in the control and we therefore did not have to adjust data using the Abott's formula. Efficacy of WHO papers was confirmed before and after field work, and 100% mortality was achieved using SUA (Susceptible colony).
Species identification
Field-collected adult A. gambiae complex mosquitoes were identified to species level using the polymerase chain reaction (PCR) assay of Scott et al. (1993) (n = 360). A. gambiae sensu stricto (s.s) molecular form identification (M or S) was performed using the PCR assay described by Favia et al. (1997). DNA was extracted using the salt-based method of Collins et al. (1987).
Sporozoite detection
The heads and thoraxes of individual wild-caught female mosquitoes (n = 187) were tested for the presence of P. falciparum sporozoites using an enzyme-linked immunosorbent assay (ELISA) (Wirtz et al. 1987). A positive control (recombinant P. falciparum) and a negative control (uninfected female A. gambiae s.s.) were included. Results were analyzed photometrically at 405 nm (Ascent Multiskan RC vl. 5.0, Genesis version 3.03, Labsystems). All positive samples were retested using the above-mentioned ELISA assay to confirm infection.
Resistance mechanisms
Hydrolysis probe/TaqMan® and restriction fragment-length polymorphism analysis for target-site resistance
Primers used in the hydrolysis probe real-time experiments were supplied by Inqaba Biotechnical Industries (Hatfield) and probes were supplied by Applied Biosystems, Inc. PCR for hydrolysis probe analysis was performed using a CFX96™ Real-Time system (Bio-Rad).
Rdl PCR
The Rdl assay (Bass et al. 2008) was used to detect the Rdl mutation associated with dieldrin resistance (n = 43). Rdl homozygous (RR and SS) and heterozygous (RS) positive controls as well as no template controls were included in all assays. Genotypes were assigned to samples based on allelic discrimination software (CFX96 Real-Time system [Bio-Rad]).
ace-1R or G119S
The restriction fragment-length polymorphism PCR described by Weill et al. (2004) was used to detect the ace-1R resistance mutation in the acetylcholinesterase gene. DNA was extracted from specimens (n = 15) that survived exposure to 0.1% bendiocarb as well as from specimens that died after exposure. The Alu1 restriction enzyme (Roche Diagnostics) digest was performed to determine if the survivors had the ace-1R mutation associated with carbamate resistance. The ace-1R mutation was identified by the presence of a 120 bp and 74 bp DNA fragment, respectively, on ethidium bromide-stained 2.5% TAE agarose gels. Wild-type specimens are recognized by the presence of a 194 bp fragment. Specimens representative of the wild-type sample (SS), heterozygous-resistant (RS), and homozygous-resistant (RR) genotypes, as confirmed by sequencing, were included as positive controls.
Kdr PCR
This assay was used to detect the kdr (L1014F and L1014S) mutations associated with DDT and pyrethroid resistance. The protocol of Bass et al. (2007) was used. Kdr homozygous (RR and SS) and heterozygous (RS) positive controls as well as no template controls were included in all assays. A total of 100 specimens that were exposed to deltamethrin were analyzed. Genotypes were assigned to samples based on allelic discrimination software (CFX96 Real-Time system [Bio-Rad]).
Synergist assays for the determination of metabolic enzyme involvement
Synergist assays were performed by exposing samples drawn from the COGS colony to either 4% piperonyl butoxide (PBO), an inhibitor of monooxygenases, 20% diethyl maleate, an inhibitor of glutathione-S-transferases (GSTs), or 20% triphenyl phosphate (TPP), an esterase inhibitor for 1 h. Synergized samples were then exposed concurrently with nonsynergized samples (drawn from the same cohort of colony mosquitoes) to either 4% DDT or 0.05% deltamethrin for 1 h. Controls included concurrent exposure of samples to untreated papers as well as exposure of samples to synergists without subsequent exposure to insecticide (Brogdon et al. 1990, 1997, Chareonviriyapha et al. 2003). Final mortality 24 h postexposure was compared between synergized and unsynergized samples using the two-sample t-test (Statistix 7- Analytical Software), following confirmation of zero mortality in any of the controls. A significance level of 5% was set for all t-test analysis.
Biochemical analysis for the detection for increased metabolic enzyme activity
Specimens for biochemical analysis were flash-frozen and transferred to 96-well microtiter plates. Each specimen was homogenized in 200 μl distilled water and assayed for protein content according to the method of Bradford, general alpha (α)- and beta (β)-esterase activity, altered acetylcholinesterase activity (Penilla et al. 1998), cytochrome P450 activity (Brogdon et al. 1997), and GST activity (Habig et al. 1974).
Three-day-old nonblood-fed adult female mosquitoes were used for biochemical analysis. To test for an association between the insecticide-resistant phenotype and enzyme levels, samples of presumed insecticide-resistant COGS (based on bioassay data for this colony) (n = 144) were concurrently assayed with insecticide-susceptible samples from an A. gambiae s.s.–susceptible colony (SUA). Mean enzyme activities were compared between COGS and SUA using two-sample t-tests (Statistix 7- Analytical Software). A significance level of 5% was set for all t-test analysis.
Microarray experimental procedures to identify specific transcripts involved in metabolic enzyme detoxification
Three-day-old COGS females were exposed for 1 h to 0.05% deltamethrin treated papers (WHO 1998). Postexposure knocked down (moribund) adults were collected as the susceptible sample, and those females that were still actively flying were collected as the resistant sample. Gene transcription levels in resistant females were compared to those from the susceptible samples. All comparisons consisted of three biological and two technical repeats. Dye swaps were also performed for each set of experiments to eliminate dye bias. Total RNA was extracted and RNA prepared and hybridized as described by David et al. (2005). The microarray slides were scanned using a Genepix 4000B scanner (Molecular Devices) until a green/red dye ratio of 1/1 was obtained. The quality of spots and background intensities were examined. The raw intensity values were analyzed with Limma software package (version 2.12.0) (Smyth 2005), run in the program R (version 2.8.0) (
Results
Baseline data
A total of 523 A. gambiae complex specimens were collected. Four hundred thirteen specimens were used for WHO insecticide susceptibility tests in the field. Full susceptibility to one insecticide (malathion) and resistance to the other four insecticides (dieldrin, bendiocarb, DDT, and deltamethrin) was recorded (Table 1). One hundred ten females were transported live to Johannesburg for additional laboratory studies on their F1 progeny.
Species identification was performed on 360/523 A. gambiae complex mosquitoes (68%) and all were identified as A. gambiae s.s. As no other species were identified, it was assumed that the remaining 32% were also A. gambiae s.s. Specimens were further analyzed to determine the presence of the two molecular forms and 95.4% of the identified A. gambiae s.s. were molecularly characterized as the S-form. One specimen (0.3%) was identified as M-form and one specimen (0.3%) was identified as a hybrid M/S. The remaining 4% could not be typed to either M or S-form.
The 110 wild-caught females reserved for additional laboratory analysis, as well as PCR-identified specimens (n = 77) preserved after field insecticide susceptibility testing, were assayed to determine the percentage infected with P. falciparum sporozoites. The P. falciparum infection rate was determined to be 9.63%.
Characterization of resistance
Target site resistance: dieldrin resistance (Rdl)
Forty-three specimens that were exposed to dieldrin were molecularly assayed for the Rdl-mutation. All the specimens that survived dieldrin exposure (n = 30) and were successfully amplified were either RR or RS genotypes (Table 2). Those mosquitoes that were susceptible to dieldrin (n = 13) carried the SS genotype. The genotype of 10/43 A. gambiae could not be assigned (Table 2).
SS, susceptible wild type; RS, heterozygous for the Rdl mutation; RR, homozygous resistant for the Rdl mutation.
Bendiocarb resistance (ace-1R )
Bendiocarb resistance was extremely low in the field population and quickly disappeared from the COGS colony (Table 1). For this reason, it was not possible to investigate the potential metabolic mechanisms that might be involved. However, specimens were analyzed for the presence of the ace-1R target site mutation. The four survivors from the bendiocarb exposure (Table 1), and 11 mosquitoes that died after exposure (randomly selected from 104 samples) were analyzed and all showed the wild-type (SS) genotype.
DDT and pyrethroid resistance
The A. gambiae s.s. population exposed in the field was 100% resistant to DDT (Table 1). F1 A. gambiae s.s. progeny from wild-caught females identified as S molecular form were pooled to establish the COGS laboratory colony, allowing for further characterization of DDT and pyrethroid resistance. This colony remained 100% resistant to DDT even after 11 generations of laboratory rearing with no exposure to DDT to exert selective pressure. Continuous exposure for 24 h on 4% DDT resulted in a mortality of only 60% (n = 100).
Pyrethroid resistance remained high in the COGS colony after 11 unselected generations. Resistance to three different pyrethroids, deltamethrin (48% mortality), permethrin (20%), and lambda-cyhalothrin (31%), was recorded. Resistance to pyrethroids and DDT serves as an indicator of target site resistance (knockdown resistance kdr), although other mechanisms, such as metabolic resistance, could also be involved (Brooke 2008). Both possibilities were investigated.
Knockdown resistance (kdr)
The presence of the L1014S (kdr-e) and the L1014F (kdr-w) mutations were investigated using the hydrolysis probe or TaqMan assay developed by Bass et al. (2007). All specimens exposed to deltamethrin (n = 100) were analyzed to determine the presence of these mutations in the Pointe Noire population. No wild-type (SS) genotypes were identified and all the specimens carried either the kdr-e or the kdr-w mutations or both (Table 3).
Carried out with hydrolysis probe analysis.
Performed without subsequent sequence analysis. No Leu-Leu genotype was found.
Genotype frequency as a proportion.
Ser, L1014S (kdr-e); Phe, L1014F (kdr-w); Leu, wild type.
To confirm the kdr results, 52/100 specimens were also sequenced. Gene frequencies were calculated for pyrethroid resistant and susceptible samples. There were no significant differences in L1014S (X2 = 2.43; df = 1; p = 0.12) and L1014F (X2 = 3.38; df = 1; p = 0.07) frequencies between the resistant and susceptible cohorts based on two-by-two contingency tables. However, analysis of genotype proportions showed significant variation in the assortment of genotypes between the resistant and susceptible cohorts (X2 = 7.08, df = 2; p = 0.03). This variation is predominantly accounted for by a significantly higher proportion of L1014F homozygotes in the resistant cohort (X2 = 7.42; df = 1; p = 0.01 based on two-by-two contingency tables) (Table 3).
Metabolic resistance: As metabolic resistance is associated with the increased activity of specific enzyme groups (P450s, monooxygenases, GSTs, or esterases), the role of these enzyme classes in DDT and pyrethroid resistance was investigated using synergists, biochemical analysis, and microarray analysis.
Synergists
DDT resistance remained at 100% survival. Neither PBO, diethyl maleate, or TPP showed any synergistic effect when combined with DDT. Deltamethrin resistance was significantly reduced using both PBO and TPP, implicating monooxygenases and esterases, respectively, in the mediation of pyrethroid resistance (Table 4).
syn, synergized sample; unsyn, unsynergized sample.
Biochemical analysis
Due to the unavailability of a susceptible strain from the same geographical area as COGS, a susceptible A. gambiae S-form laboratory strain, SUA, was used. Even though COGS showed an increase in P450 enzyme activity than the susceptible SUA strain, this increased rate was not significant (two sample t-test: t = 1.45; df = 286; p = 0.15) (Fig. 1). There was no evidence of an altered acetylcholinesterase mechanism in COGS. General α-esterase activity was also slightly elevated when compared to SUA, but again the difference was not significant (two sample t-test: t = −0.73; df = 286; p = 0.47). However, a significantly higher level of β-esterase activity (two sample t-test: t = −6.15; df = 278; p < 0.01) was observed. A significantly higher level of GST activity was observed in the COGS colony compared to the susceptible SUA colony (two sample t-test: t = 5.8; df = 526; p < 0.01) (Fig. 1).
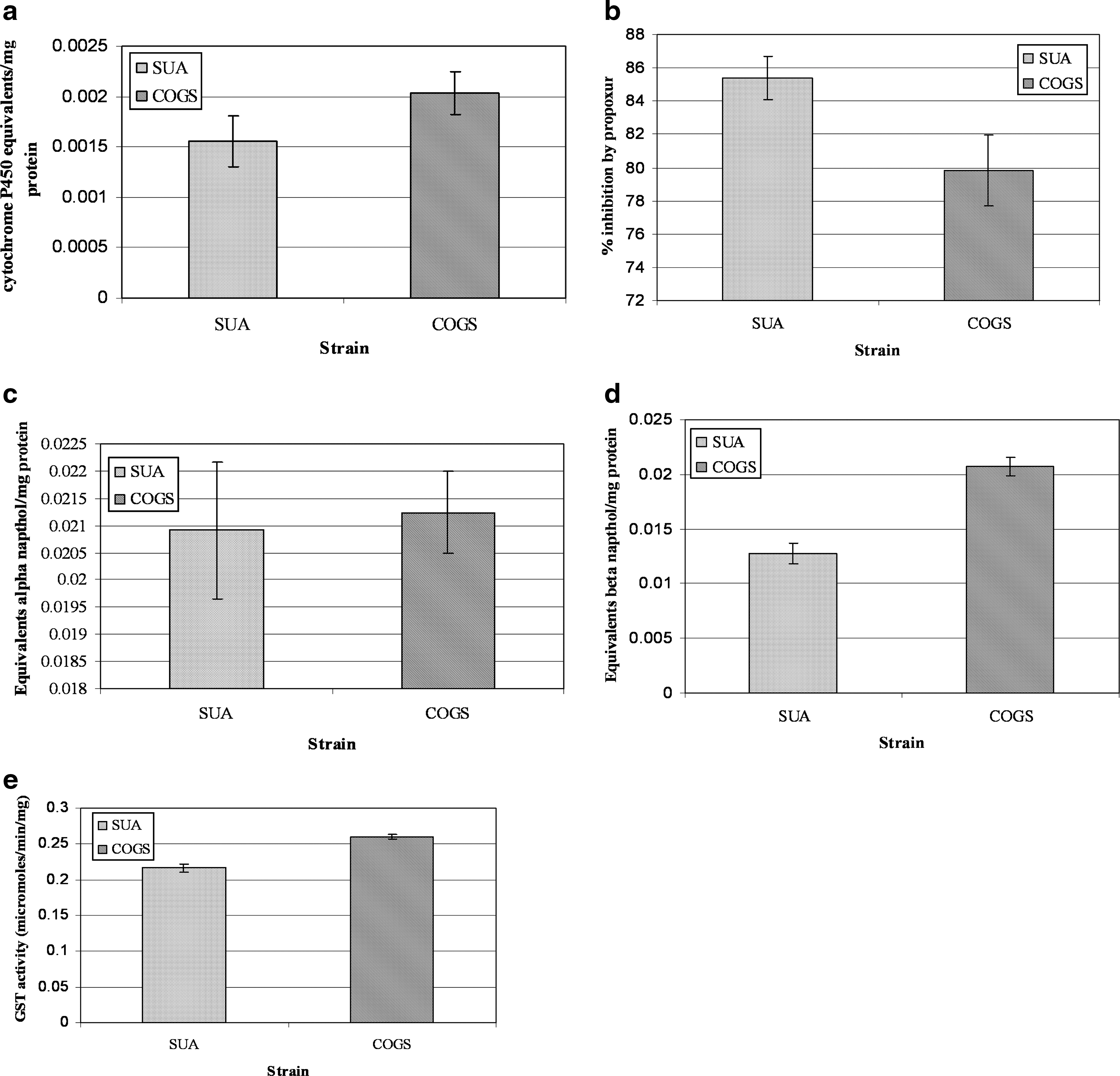
Biochemical analysis.
Microarray analysis
Of the 254 gene probes on the A. gambiae detox chip, no genes were found to be significantly differentially expressed between the resistant and susceptible specimens. Therefore, the significance level was reduced to a value of 0.01 and the fold-change cut-off was kept at 1.5 to assess if there were any overexpressed transcripts. Three genes, CYP6M2 (2.23-fold), COEJHE5E (1.63-fold), and CYP6AK1 (1.63-fold), were differentially expressed (Table 5). The highest fold-change was obtained for the P450 gene CYP6M2. As the microarray data supported synergist data, it was included in the analysis.
Discussion
Pointe Noire, as in Cameroon and Ghana, showed an abundance of S-form A. gambiae with a high P. falciparum infection rate (Coetzee et al. 2006, Etang et al. 2009, de Souza et al. 2010). Multiple insecticide resistance was observed in this population, which showed full susceptibility to only one of the insecticide classes tested, organophosphates.
Owing to mosquito dispersal, it is conceivable that the multiple resistances observed at this locality also occur at other localities in the Republic of the Congo and neighboring regions, although the levels of resistance are likely to vary by insecticide class. For example, Vezenegho et al. (2009) reported multiple insecticide resistance at three widely dispersed localities in Guinea Conakry. The levels of resistance between the three localities varied depending on the insecticide classes tested. Coetzee et al. (2006) and Vezenegho et al. (2009) also reported susceptibility to malathion (organophosphate) in Ghana and Guinea Conakry, respectively, suggesting that particular patterns of resistance or susceptibility to insecticides are widespread in West Africa. Similar patterns were observed when looking at P. falciparum infection rates. In the study described by Vezenegho et al. (2009), two of the three localities reported infection rates between 5% and 6%, whereas the third showed infection rates of 21%, emphasizing the similarities as well as the differences that can be observed within a given region.
The dieldrin resistance recorded can be attributed to reduced target-site sensitivity (Rdl) although a metabolic component cannot be excluded in the current study. Dieldrin is no longer used following a ban in the 1960s (De Kom and Dewan 2007), and yet the resistance phenotype has persisted without apparent selection. This phenomenon has also been observed in Ghana (Brooke et al. 2006) and in two laboratory strains of A. gambiae (Brooke et al. 2002).
Bendiocarb survival was only recorded in four of the 106 A. gambiae s.s. mosquitoes exposed (96.2% mortality). Using the WHO criteria for presence of resistance (98%–100% is considered susceptible, 80%–97% resistance is suspected, <80% resistance is confirmed) this population would be classified as possibly resistant, requiring further investigation. As DNA was extracted for species identifications, it was decided to simultaneously screen for the presence of the ace-1R mutation. The absence of this mutation in the four survivors may suggest a metabolic resistance mechanism or they may have survived for other reasons (cuticle thickness, avoidance of the treated papers, etc.). Nevertheless, susceptibility levels need to be periodically monitored if carbamates are to be considered for vector control in this area.
DDT resistance in wild mosquitoes from Pointe Noire reached a maximum of 100%, implying that the phenotype is fixed in this population. Fixation for DDT resistance was mirrored in the COGS colony, even after 11 generations without any selection pressure. Pre-exposure to synergists indicated that a metabolic system is most likely not involved in production of the resistance phenotype even though the biochemical analysis showed a significant increase in GST activity. Although GSTs have been implicated in DDT resistance in various other vector populations (Che-Mendoza et al. 2009), the increase in general GST activity in this population does not appear to be directly associated with DDT resistance.
Importantly, no kdr SS specimens were identified, and all those specimens analyzed carried one or both kdr mutations (L1014F and L1014S), indicating that DDT resistance is primarily based on reduced target-site sensitivity. However, the presence of two heterozygous (RS) specimens showed that the wild-type allele is still present at a very low frequency. Fixation for DDT resistance at the molecular level may yet occur in the Pointe Noir population if the level of insecticide pressure is sufficient.
Considering that kdr is supposed to confer cross-resistance between pyrethroids and DDT (Soderlund and Knipple 2003, Davies et al. 2007), the 26% pyrethroid survival observed here, compared with 100% DDT survival, is not easily explained by the assortment of kdr. Since DDT and pyrethroids both target the sodium-ion channel, the difference in the impact of kdr between these two insecticide classes is surprising. From the data presented, only homozygotes for the L1014F kdr mutation showed evidence of selection following pyrethroid intoxication. Yet, pyrethroid resistance is evidently far from fixation despite the near fixation of kdr genotypes. The presence of either or both kdr mutations, even in individuals scored as homozygous resistant, does not necessarily produce a pyrethroid resistant phenotype. The lack of correlation between kdr and pyrethroid resistance has previously been described by Ramphul et al. (2009).
The synergist data provide important information concerning the mechanism of pyrethroid resistance. Pre-exposure to PBO and TPP significantly increased subsequent susceptibility to deltamethrin, implying that metabolic detoxification plays a fundamental role in the pyrethroid resistance in this population. The candidate detoxification enzyme systems proposed are monooxygenases and esterases although biochemical analysis did not reveal a significant increase in P450 activity when compared to the susceptible strain, SUA. This discrepancy may be due to strain variation given that SUA and COGS do not share the same genetic background.
Initial microarray analysis did not identify any candidate resistance associated genes based on differential expression between resistant and susceptible phenotypes. However, after lowering the cut-off criteria, three potential genes associated with resistance were identified. The highest fold differentially transcribed gene was CYP6M7, providing further evidence of a monooxygenase based pyrethroid resistance mechanism. Adjusting cut-off criteria, however, is not recommended and the next aim will be to select a pyrethroid resistant strain from the current COGS colony and perform comparisons between the selected and unselected strains. These results will then be confirmed using quantitative real-time PCR.
In conclusion, A. gambiae S-form is the primary vector of malaria in Pointe Noire and it has developed resistance to multiple insecticide classes. DDT resistance is conferred by the L1014F and L1014S kdr mutations, whereas pyrethroid resistance is mainly, but not exclusively mediated by monooxygenase and esterase detoxification. The high level of DDT resistance obviously negates this insecticide for use in a malaria vector control in the Pointe Noire region. Pyrethroid resistance remains high in the laboratory-reared colony with high levels of resistance recorded between the different chemicals; however, deltamethrin gave the highest mortality (48%). As only pyrethroids are used for treating bed nets, it would be advantageous in terms of resistance management to use either organophosphates or carbamates for indoor residual house spraying. The geographical range of the multiple insecticide resistance observed in this study needs to be investigated.
Footnotes
Acknowledgments
Financial support was provided by the National Health Laboratory Service Research Trust and the South African National Research foundation to LLK and the South African DST/NRF Research Chair grant to MC. We are grateful to Dr. C. Bass for supplying positive controls used in this study as well as for valuable comments from reviewers. The authors thank Dr. H. Ranson for providing the “detox chip” used in this study.
Disclosure Statement
The authors declare no conflict of interest.