
Abstract
Lyme borreliosis is an arthropod-borne disease transmitted by the Ixodes tick. This spirochetal infection is first characterized by a local cutaneous inflammation, the erythema migrans. The skin constitutes a key interface in the development of the disease. During Borrelia inoculation, tick saliva affects the innate and adaptive immunity of the vertebrate host skin. Some key mediators of innate immunity such as antimicrobial peptides (cathelicidin and defensin families) have been identified as important initiators of skin inflammation. We analyzed the role of tick saliva on integumental innate immunity using different protocols of Borrelia infection, via syringe or direct tick transmission. When syringe inoculation was used, Borrelia triggered skin inflammation with induction of CRAMP, the mouse cathelicidin, and tumor necrosis factor-alpha. However, when Borrelia was transmitted directly via the tick, we observed a significant repression of inflammatory genes, suggesting a critical role of tick saliva in skin innate immunity. For all the protocols tested, a peak of intense Borrelia multiplication occurred in the skin between days 5 and 15, before bacterial dissemination to target organs. We conclude that Borrelia pathogens specifically use the tick saliva to facilitate their transmission to the host and that the skin constitutes an essential interface in the development of Lyme disease.
Introduction
Ticks help the development and multiplication of the pathogens in the host by their saliva. They challenge the host immune system through a broad variety of molecular mechanisms (Brossard and Wikel 2004, Hovius et al. 2007). The effect of tick saliva on host adaptive immunity has been documented for a long time (for a review, see Wikel 1999). Wikel and collaborators (1978) described that tick saliva downregulates in vitro the T-cell proliferation. Years later, an IL-2 binding protein in the saliva of I. scapularis was described in the suppression of T-cell proliferation (Gillespie et al. 2001). Recently, a tick saliva protein, Salp15, was identified as a potential candidate for this immunosuppressive effect (Garg et al. 2006). Tick saliva has also been described to promote the development of a Th-2 response that facilitates Borrelia transmission (Skallová et al. 2008). The effect of tick saliva on innate immunity has been studied more recently. Two proteins of I. scapularis salivary glands, Isac and Salp20, have been described as inhibitors of the alternative pathway of the complement (Valenzuela et al. 2000, Tyson et al. 2008).
In arthropod-borne diseases, the skin plays a significant role in the control of skin infections by displaying an efficient immune response (Frischknecht 2007, Krause et al. 2009). During the infectious blood meal, the tick hypostome dilacerates and penetrates the epidermis and reaches the dermis, where Borrelia is inoculated. This induces a local immune response constituted by the secretion of chemokines, cytokines, and antimicrobial molecules (antimicrobial peptides [AMPs]). AMPs belonging to the cathelicidin and defensin gene families are of particular importance to the mammalian skin innate defense system (Braff and Gallo 2006). In mouse models, the cathelicidin CRAMP is an important native component of innate host defense by providing protection against necrotic skin infection caused by Group A Streptococci (Nizet et al. 2001). Similarly, defensin has been shown to control bacteria in skin infections (Schröder and Harder 2006). So far, the potential role of AMPs has never been investigated in vivo in the context of arthropod-borne diseases. Recently, we demonstrated in an in vitro model that defensin and cathelicidin are secreted by resident skin cells incubated with Borrelia and inhibited in the presence of tick saliva (Marchal et al. 2009, 2011). Here, experiments were carried out with C3H/HeN mice to assess the role of these molecules in vivo in the transmission of B. burgdorferi ss and the specificity of their induction. We also explored the role of tick saliva on Borrelia multiplication in the skin and on the dissemination.
Materials and Methods
C3H/HeN mice were purchased from Charles River Laboratories. B. burgdorferi ss N40 strain was cultured in BSK-H complete medium (Sigma) at 33°C and used at low passage for mouse infection. For nymph infection, Ixodes ricinus larvae were allowed to feed to repletion on C3H/HeN mouse previously intradermally infected via syringe inoculation with 105 B. burgdorferi ss N40. After 3–5 days, replete larvae were kept in a humidified chamber until they molt into nymphs. They were checked for the presence of B. burgdorferi ss by quantitative polymerase chain reaction (PCR).
Five different infection protocols of mice were considered in the kinetic study. (A) In the tick transmission model, five B. burgdorferi-infected nymphs per mouse were fed according to Mbow et al. (1994). (B) In the syringe transmission model, mice were inoculated intradermally in the dorsal thoracic area with 103 B. burgdorferi/100 μL. (C) Five uninfected nymphs were allowed to feed to repletion on each mouse, and then at day 5 when all nymphs detached, 100 μL of B. burgdorferi (103 bacteria) were inoculated at the site of the tick bites as described in protocol B. (D) Mice, just bitten by uninfected nymphs, were directly used to measure the skin inflammation induced by the tick bite itself. (E) As a positive control for the induction of AMPs, mice were infected subcutaneously with an invasive human Group A Streptococcus strain as described by Nizet et al. (2001). For these five protocols, at the site of needle inoculation or tick bite, skin biopsies were collected at 5 and 24 h and at days 3, 5, 7, 15, and 30 postinfection (p.i.) and frozen at −80°C either directly for Borrelia quantification by PCR or in TRI reagent (Invitrogen). Infected ticks were collected at each time point of the kinetics and checked by PCR for the presence of Borrelia.
Mouse sera were collected at each time point of the kinetics and IgG antibody titers for B. burgdorferi were determined by enzyme-linked immunosorbent assay as previously described (Soulas et al. 2005).
At the different time points p.i., specific organs from each mouse were dissected aseptically. For B. burgdorferi detection by culture, dissected heart, joint, and ear tissue were transferred in BSK-H medium at 33°C and examined weekly by dark-field microscopy for the presence of spirochetes. For B. burgdorferi detection by quantitative PCR, DNA was extracted from the organs of individual mice (bladder and joint) on a MagNA Pure apparatus as described by the manufacturer (Roche Diagnostics). For Borrelia quantification in the skin, mice were inoculated intradermally with different bacterial concentrations (10, 102, 103, or 104 Borrelia) in 20 μL for the quantification at day 0 and in 100 μL for the kinetic study. The skin was collected with a 3-mm-diameter biopsy punch (Lab Stiefel®) at the site of the inoculation for Borrelia quantification. Qualitative amplification targeting the fla gene was carried out as described by Jaulhac et al. (2000). Quantification of the B. burgdorferi-specific fla gene was performed on a LightCycler system (Roche Diagnostics) as previously described (Woods et al. 2008). Quantification of the mouse-specific gapdh gene was done on an ABI Prism 7000 instrument (Applied Biosystem), using a commercial kit (TaqMan rodent GADPH control reagent; Applied Biosystem). Number of B. burgdorferi spirochetes in tissue specimens was normalized to 104 gapdh DNA copies.
Total RNA was extracted from 10 mg of mouse skin using TRI reagent according to the manufacturer (Invitrogen). Samples were treated by DNAse (Ambion) and first-strand cDNA was synthesized from 2 μg of total RNA using SuperScript II reverse transcriptase (Invitrogen Life Technologies). GAPDH quantification was performed as an internal standard. Relative expression levels were calculated using an uninfected animal as a calibrator. Amplification and detection of mRNA from target genes were performed with an ABI 7000 system with the following thermal profile: 95°C for 10 min, 50 cycles of 95°C for 15 s, 60°C for 1 min. Primers used are listed in Table 1.
Each experiment was carried out at least three times. For each RT-PCR on the skin, at least two extractions were done for each mouse in each experiment.
Results
We first analyzed Borrelia transmission according to the different protocols tested. At each time point of the infection with five infected nymphs per mouse, ticks were collected and checked by PCR. They were positive at 80% for Borrelia (Table 2-IA). At 24 h, all infected ticks were fixed on mouse skin and they were all detached at day 5. Whatever the protocols of infection, Borrelia were first detected in the skin at the inoculation site by PCR: at day 7 in mice infected with Borrelia via infected nymphs and at day 3 in syringe-infected mice (Table 2). The presence of tick saliva in the skin delayed the detection of Borrelia because PCR detection was positive at day 5 only (Table 2-IC). The serology turned out to be positive at day 15 whatever the protocols used.
(I) Measure of Borrelia burgdorferi transmission. (II) Measure of B. burgdorferi dissemination by in vitro culture of infected organs “C” or by PCR. (A) Borrelia-infected nymphs; (B) syringe inoculation of Borrelia; (C) bite of uninfected nymphs followed by syringe transmission of Borrelia. The presence of Borrelia in the skin and in the infected nymphal ticks was measured by PCR.
PCR, polymerase chain reaction; ND, not done.
Then, we measured the effect of tick saliva on Borrelia dissemination to distant organs (Table 2-II). We tested the bladder, joint, and heart by culture and/or quantitative PCR for the presence of Borrelia. When infected nymphs were used to analyze the kinetic of Borrelia transmission, all distant organs were positive by quantitative PCR at day 15 only (Table 2-IIA). Transmission of Borrelia by syringe inoculation enhanced the speed of dissemination because the joint was already positive at day 5 and the heart at day 7 (Table 2-IIB). The presence of tick saliva in the skin before Borrelia inoculation delayed the dissemination because the heart was positive by culture at day 15 versus day 7 in the absence of tick saliva (Table 2-IIC).
In C57/BL6 mice, cathelicidin has been previously shown to be a key element in the control of skin infection induced by group A Streptococcus (Nizet et al. 2001). We tested whether this AMP and those of the defensin family might play a role during the transmission of Borrelia to the vertebrate host. We observed that the bite of infected nymphs induced a slight inflammatory response in the skin, with a peak of mBD14 and mBD3 defensins at days 3 and 5 pi, respectively (Fig. 1A). Interestingly, in the absence of tick saliva, the needle transmission of Borrelia induced a significant increase of cathelicidin (CRAMP) and tumor necrosis factor-alpha (TNF-α), with a peak at 24 h and at day 7, respectively (Fig. 1B). To investigate more in details the effect of tick bite and saliva on skin inflammation, we first analyzed mouse skin infested by uninfected nymphs followed by Borrelia syringe inoculation (Fig. 1C). We let uninfected ticks feed to repletion, and after their complete detachment at day 5, we injected Borrelia as in Figure 1B. We observed that the presence of tick saliva in the skin strongly inhibited the inflammatory response. MCP1 was the only inflammatory gene induced at 5 and 24 h and at day 7. No CRAMP or TNF induction was detected. The bite of uninfected nymphs by itself did not induce a significant level of the inflammatory genes tested (Fig. 1D). The effect was specific of Borrelia injection because the inoculation of the BSK medium did not induce inflammatory genes (data not shown) and the inflammatory profile induced by Streptococcus group A injection with a similar kinetic protocol exhibited a different inflammatory profile (Fig. 1E). Indeed, a very strong induction of the mBD3 defensin was observed in the latter protocol with a peak at day 3 and CRAMP reached a level similar to the level induced by Borrelia with a 50-fold increase at day 3.
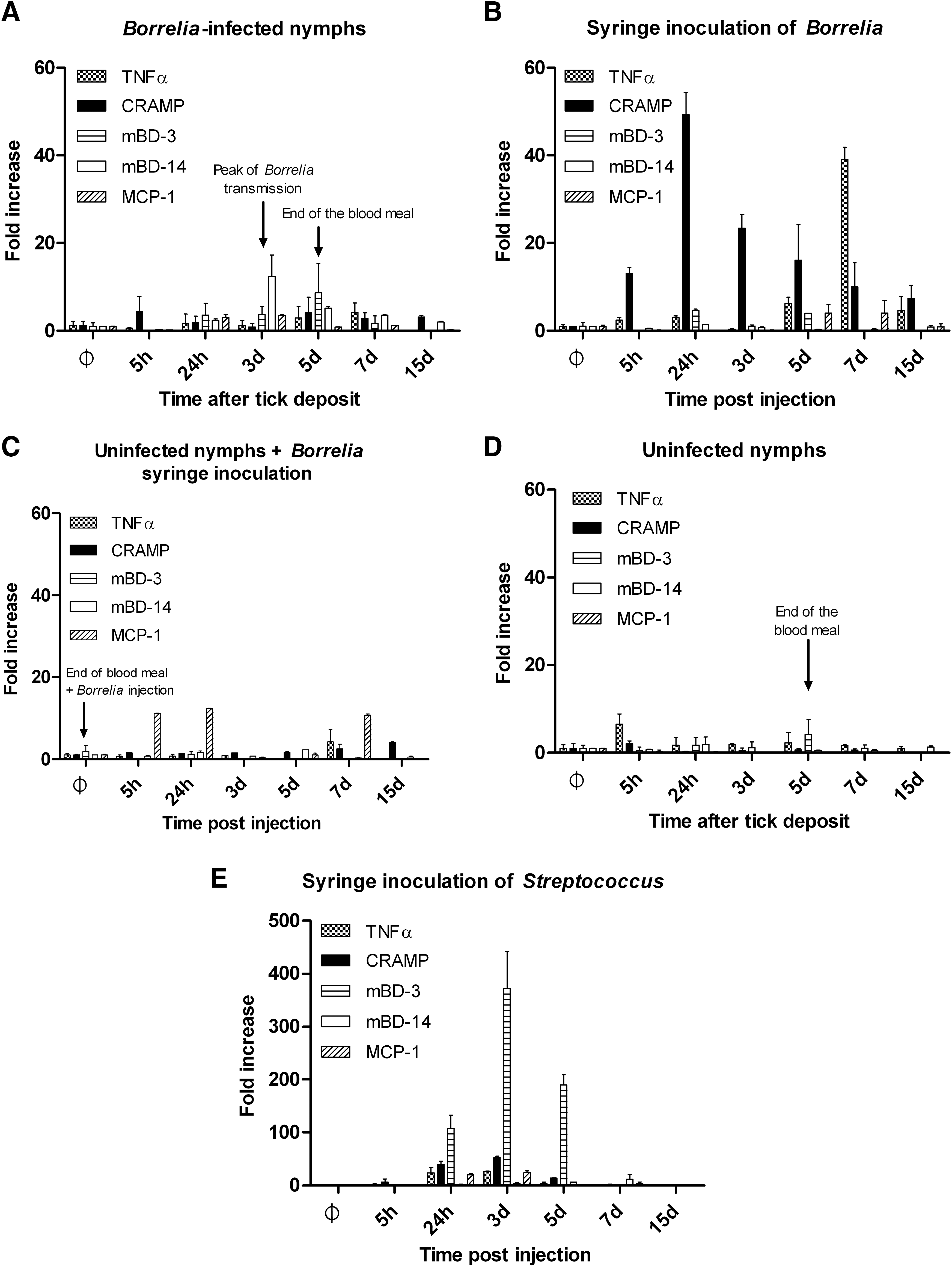
Tick saliva has an immunosuppressive effect on the induction of mouse skin inflammation. Measure of inflammatory genes by RT-PCR at different time points after infection: cathelicidin (CRAMP), defensins (mBD3 and mBD14), tumor necrosis factor-alpha, and the chemokine, MCP-1.
We then made a quantification of Borrelia by PCR at the site of inoculation at the different time points of the kinetic study. Each protocol of infection exhibited a peak of significant Borrelia multiplication in the skin of infected mice: day 5 for syringe inoculation and day 15 for nymph transmission. Interestingly, the presence of tick saliva in the skin delayed and decreased the peak of Borrelia multiplication at day 7 (Fig. 2). The bacterial load detected after a natural tick infection was less important than in the two other infection protocols likely because of the much smaller amount of Borrelia inoculated via infected ticks.

Borrelia intensively multiplies in the skin at different time points after infection according to the protocol of infection. Quantification by PCR of B. burgdorferi at the site of inoculation at different time points after the beginning of infection. Three protocols of Borrelia infection were used: through infected nymphs, through syringe inoculation, and through the bite of uninfected nymphs followed by needle transmission of Borrelia.
By quantitative PCR, we tried to measure the threshold of Borrelia detection in the skin according to the dose and the time pi. We quantified by PCR the spirochete load in the skin after inoculation of different numbers of Borrelia: 10, 102, 103, and 104. It was necessary to inoculate 102 spirochetes to be able to detect the bacteria in the skin at day 0 at the inoculation site, with 0.002 flagellin copies per 104 murine Gapdh (Fig. 3A). At 103 Borrelia, an average of 0.07/fla was detected and 1.24 with 104 Borrelia. Then, we made a kinetic study with different Borrelia concentrations and collected skin biopsies at days 1, 3, 7, and 15. An inoculum of 10 Borrelia was sufficient to allow detection in the skin at day 7, whereas a 103 inoculum was already detected at day 3, with 0.07 flagellin copies per 104 murine Gapdh (Fig. 3B). The inoculum of 104 showed a peculiar profile: a decrease at 24 h and then a regular increase until day 15 that could correspond to congregate of spirochetes in tissues (Hodzic et al. 2003).
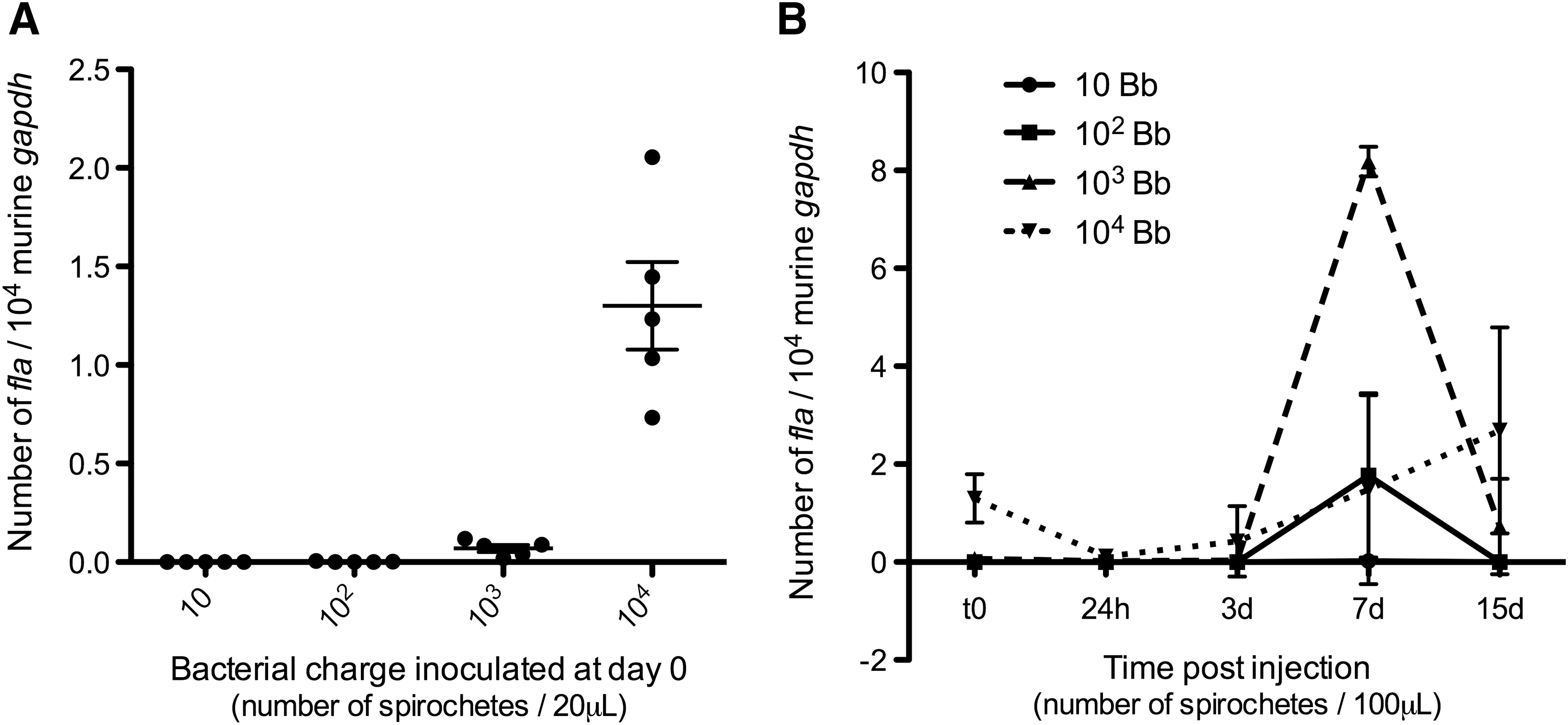
Threshold of Borrelia detection in the skin.
A comparison of the inflammatory profile for these different inocula demonstrated a dose dependence of inflammation (Fig. 4) and of dissemination (supplemental data). The 102 Borrelia induced a peak of TNF at day 7 corresponding to the peak of Borrelia; a similar profile was observed for 103 Borrelia (Fig. 4A, B). However, AMPs were not significantly induced at 102. The dose of 104 Borrelia induced a peak of mBD3 defensin at 24 h, some CRAMP, and a peak of TNF at day 15 (Fig. 4C).
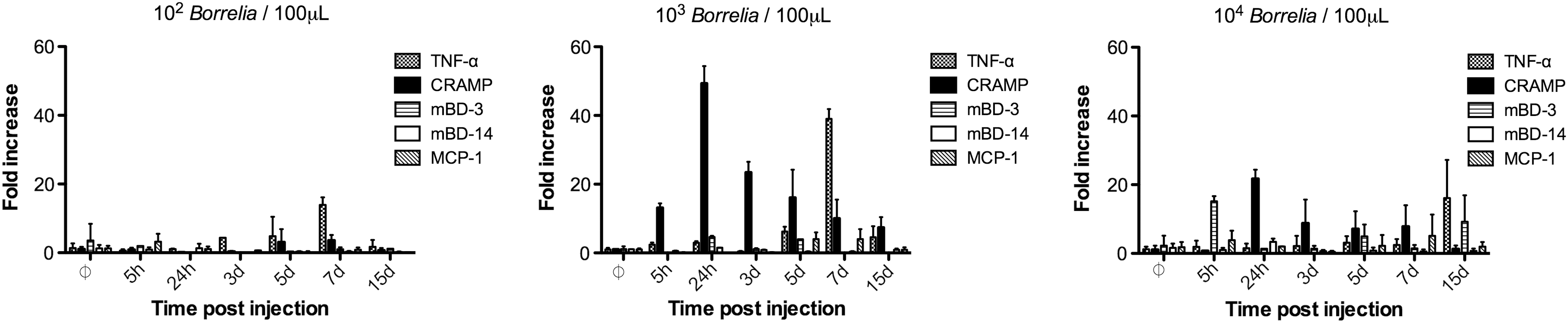
Inflammatory profile of the skin of mice inoculated via syringe with different concentrations of Borrelia: 102, 103, 104.
Discussion
Pathogens transmitted via arthropods have evolved numerous strategies to allow their transmission to the vertebrate host, using the potent anti-inflammatory and immunomodulatory effect of arthropod saliva. In Lyme disease, the tick is essential for Borrelia: it significantly enhances its virulence by its saliva (Pechová et al. 2002, Hovius et al. 2008). It was recently shown that the B. burgdorferi ss N40 isolate with a high passage that was no longer able to infect C3H mice recovered its pathogenesis in mice when it was inoculated via I. scapularis (Adusumilli et al. 2010). In tick saliva, numerous molecules have been identified with immunosuppressive effects (Hovius et al. 2007). So far, the tick saliva was never studied as a potential immunosuppressor of skin AMPs. Defensins and cathelicidins are described as important actors in different skin inflammations (Dorschner et al. 2001, Ong et al. 2002, Braff and Gallo 2006). They could be particularly relevant in the transmission of Lyme disease because the tick stays attached to the skin of its host for several days before an erythema migrans develops there and constitutes the first clinical symptom of the disease (Steere et al. 2004). Group A Streptococcus and Borrelia induced a different inflammatory profile underlining that skin innate immunity is specific and confirming previous observations (Dinulos et al. 2003). It is noteworthy that, when the tick saliva was tested in vitro for its role on Borrelia inflammation, it exhibits a strong immunosuppressive effect on skin AMPs (Marchal et al. 2009, 2011). We described here that tick saliva also impacts in vivo innate immunity by inhibiting AMP secretion. Identification of tick saliva proteins with an immunosuppressive effect on skin innate immunity, especially affecting AMP, could be part of an effective vaccine against tick-borne pathogens (Wikel 1999, Schuijt et al. 2011). Interestingly, MCP-1 was only induced when uninfected nymphs were let to feed to repletion before syringe inoculation of Borrelia. This chemokine has been detected in erythema migrans (Zhao et al. 2007) and recently in the skin of Borrelia-infected mice (Antonara et al. 2010). It could facilitate pathogen dissemination by modifying vascular permeability as shown in dengue (Lee et al. 2006).
In Lyme disease, the transmission does not take place at the beginning of the bite because a delay is necessary for Borrelia to migrate from the midgut to the salivary glands. In North America, with nymphal I. scapularis infected with B. burgdorferi ss, it can occur after 24 h in hamster, whereas it is observed after 48 h for mice (Piesman et al. 1987, Hojgaard et al. 2008). In Europe, nymphal I. ricinus infected with B. afzelii are able to transmit the infection in <24 h after attachment (Crippa et al. 2002). The number of pathogens inoculated by Ixodes tick is difficult to evaluate because the infective blood meal lasts for several days. Comparing Figures 2 and 3A quantifications, we can estimate that around a hundred of Borrelia is inoculated by an infected tick. Interestingly, in our study, according to the syringe-inoculated dose, it seems that a too high dose of Borrelia (104) disrupts the skin immune response. This could be linked to a too high number of bacteria inoculated locally, which finally would be detrimental to Borrelia survival at first. With such an inoculum, we unexpectedly observed first a decrease of bacterial number and then a regular increase. In addition, we showed that whatever the inoculation protocol tested, a bacterial multiplication occurs in the skin as previously described (Hodzic et al. 2003). Although a significant multiplication occurs in the tick midgut, the number of Borrelia in salivary glands seems to be quite low (Piesman et al. 2001). The multiplication in the skin could facilitate Borrelia infection and dissemination, pointing out that the skin plays a role of amplification site. Ten Borrelia were sufficient in our study to induce an infection as we were able to detect the spirochetes in the skin. In the kinetic studies wherein tick saliva was present together with Borrelia or just before the inoculation, we observed a longer persistence of the bacteria in the skin. It supports the fact that through tick bites, Borrelia persists longer in mice than when spirochetes are syringe inoculated (Zeidner et al. 2001).
We conclude that the skin is a key interface to initiate arthropod-borne diseases. A recent study performed with mice, syringe-inoculated with an OspC mutant of B. burgdorferi ss, exhibited the potent role of OspC to initiate skin inflammation (Antonara et al. 2010). In malaria, similar observations have been made. By imaging microscopy, it has been shown that part of the sporozoites stays in the skin to achieve a partial development (Amino et al. 2006, Gueirard et al. 2010). A better understanding of skin immunity should help to develop efficient strategies to control these diseases (Hovius et al. 2008, Sinnis and Zavala 2008).
Footnotes
Acknowledgments
The authors thank the Direction Générale de l'Armement and the Conseil Régional d'Alsace for the Ph.D. financial support to A. Kern. The authors also thank Prof. B. Betschart (Institute of Biology, Neuchâtel, Switzerland) for help in the maintenance of tick colony in this study, Chantal Delena from the French National Reference Centre of Borrelia for culture of Borrelia strains, and the REID (Réseau des Interactions Durables-Groupe tique) and the GEBLY (Groupe d'Etude sur la Borréliose de Lyme) for their stimulating discussions.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.