
Abstract
Temperature is a key factor influencing mosquito growth and development and is also known to affect insecticide efficacy. We evaluated the effects of larval rearing temperature and exposure to insecticides on adult mosquito fitness and competence for arboviral infection using Sindbis virus (SINV). We exposed newly hatched larvae of Aedes aegypti to an environmentally realistic level of insecticide malathion at 20°C and 30°C and allowed the resulting adults to feed on SINV-infected blood meal. Exposure to malathion significantly reduced survival to adulthood. Statistically significant interactions between temperature and malathion were observed for body size, estimated population growth, and SINV infection and dissemination. Malathion-exposed Ae. aegypti cohorts had significantly higher population growth at 20°C than at 30°C. Body size decreased with higher temperature and malathion-exposed females were larger than unexposed females at 20°C but not at 30°C. Viral infection and dissemination increased with larval rearing temperature and were higher in malathion-exposed than unexposed females at 30°C but not at 20°C. These results show that environmental factors, including those factors used in controlling mosquitoes, experienced by immature stages have latent effects that continue to adulthood and alter vector competence to arboviruses.
Introduction
In nature, mosquito larvae are exposed to pesticides in the presence of aforementioned stressors, but standard toxicological studies are usually conducted in the absence of these stressors. This design is likely to underestimate the lethal and chronic effects of pesticides on mosquitoes because combined effects of pesticides and environmental stressors have been shown to generate nonadditive effects (Sih et al. 2004) in many vertebrate and invertebrate taxa (Lohner and Fisher 1990, Boone and Bridges 1999, Linke-Gamenick et al. 1999, Boone and Semlitsch 2002, Liess 2002, Relyea 2003, Beketov and Liess 2006, Campero et al. 2007, Muturi et al. 2010). Pesticides can reduce the fitness of an organism by causing structural and functional alterations to the immune system (Blakley et al. 1999a, 1999b) or by modifying life history traits through reduced food intake (Ribeiro et al. 2001), diversion of energy to maintenance, and/or modification of the level and activity of target biomolecules (e.g., inhibition of enzyme activity, Campero et al. 2007). Mortality due to pesticide in the immature stages may also be associated with improved performance of the surviving individuals, especially when resources are limiting (Muturi et al. 2010). These effects may allow for the possibility of enhancement or suppression of vector competence depending on the underlying environmental conditions.
Temperature is a key determinant of vector-borne disease transmission, which may modify the effect of a pesticide on adult mosquito fitness and vector competence in several ways. First, temperature significantly influences mosquito growth and development (Alto and Juliano 2001, Lounibos et al. 2002, Bayoh and Lindsay 2004) as well as pathogen replication (Mourya et al. 2004, Westbrook et al. 2010). Second, thermal stress has been shown to induce expression of proteins that confer mosquito tolerance to some insecticides (e.g., Aedes aegypti L. and Anopheles stephensi Liston) (Patil et al. 1996). Third, temperature affects insecticide efficacy so that some chemicals become more or less lethal as temperature increases (Mayer and Ellersieck 1986). Thus, environmental temperature may facilitate pesticide-induced mortality of mosquitoes or lead to more rapid pesticide degradation, which is predicted to reduce mortality because of shorter exposure periods (Starner et al. 1999).
The present study tested the hypothesis that temperature modifies pesticide effects on adult mosquito fitness and infection with Sindbis virus (SINV, Togaviridae: Alphavirus). The arbovirus, SINV, is a single-stranded RNA virus with a wide geographic distribution circulating between Culex and Aedes species and birds (Tesh 1982). Human infections by Sindbis-related viruses produce a self-limiting febrile disease with lasting arthralgia in some instances (Niklasson and Espmark 1986, Laine et al. 2000, 2004, Kurkela et al. 2005). Several outbreaks involving human cases have occurred in South Africa, northern Europe, and Russia (Jupp et al. 1986, Lundström et al. 1991, Laine et al. 2004, Kurkela et al. 2008). Our model system included the yellow fever mosquito Ae. aegypti and insecticide malathion. Ae. aegypti is an important vector of human disease agents including dengue, yellow fever, and chikungunya viruses (Gubler 2004, Thavara et al. 2009). Malathion is used in agricultural settings and mosquito control and is readily available to the public for residential application. Malathion is applied as an ultralow-volume spray to control adult mosquitoes and may drift for long distances and contaminate other aquatic systems including container habitats (Walker 2000). Environmental concentrations of malathion up to 1 mg/L have been recorded in aquatic systems (U.S. Department of Agriculture 1997). Our aim was to use a model system to identify environmental influences on the immature stages of mosquitoes that may continue to adulthood and alter vector competence for arboviruses.
Materials and Methods
Life history traits and population growth
Mosquitoes used in this experiment were F7 progeny of Ae. aegypti originally collected in south Florida. Experimental units consisted of 4-L plastic containers with 1.6 L of filtered oak infusion (1.0-mm2 sieve), 200 larvae (∼12 h old), and 0.1 g of an equal mixture of brewer's yeast and albumin. To prepare the infusion, 500 g of oak leaves were added in 100 L of tap water and the mixture was left to steep for 3 weeks. We manipulated temperature (20°C and 30°C) and malathion (0 or 0.05 parts per million [ppm]) treatments during the immature stages in a full-factorial experiment with six replicates. Pesticide concentration was achieved by adding 3.8 mL of 20 ppm malathion to treatment replicates during the experimental setup, whereas an equal volume of deionized water was added to control replicates. A supplemental food resource of 0.2 g brewer's yeast and albumin were added on day 7. Pupae were removed daily and placed in groups of six in plastic vials until adult emergence. The adults (both males and females) were recorded, housed by treatment in paperboard cages (11 cm high × 9.5 cm diameter), and maintained on 10% sucrose at 25°C. For each replicate, mosquitoes emerging each day were placed in a new cage to create distinct ages of adult females for vector competence study (5–10 days old; see below).
For each replicate, we measured survivorship to adulthood, development time to adulthood, and female size (wing length in mm). An estimate of finite rate of increase (λ′) was also calculated for each treatment replicate:
where N o is the initial number of females (50% of initial larvae), Ax is the number of females emerging to adulthood on day x, wx is mean female size on day x, f(wx ) describes a relationship between female size and fecundity, and D is the time to oviposition (D estimated as 12) (Grill and Juliano 1996). The fecundity–size relationships (f(wx )) used to calculate λ′ was f(wx ) = 2.505 (wx 3) − 8.616, r 2 = 0.875, N = 206, and p < 0.001 (Briegel 1990). Determination of wx is based on wing length (mm) measurements of females emerging on day x.
Separate two-way ANOVAs were used to test for effects of temperature and malathion treatments on Ae. aegypti survivorship to adulthood (males + females), male development time to adulthood, and λ′. Survival data were arcsine transformed to meet the assumption of normality (SAS Institute 2002). Multivariate analysis of variance (MANOVA) was used to determine the effect of temperature and malathion on female development time and adult size. Standardized canonical coefficients were used to describe the relative contribution of life history traits to the significant treatment effects. When significant effects were obtained, pairwise differences between treatment means were compared using a Tukey–Kramer adjustment (SAS Institute 2002).
Vector competence
Females from temperature and malathion treatments were transferred to a biosafety level-2 facility and allowed to feed on SINV (MRE16 strain)-infected blood meals (Alto et al. 2005). For each blood feeding session, blood meals containing 105 plaque-forming units/mL were prepared by mixing a previously frozen SINV stock with citrated bovine blood. Female adults were starved for 24 h and then provided with 30 min access to infected blood using the Hemotek® feeding system. Fully engorged females were held at 25°C for a 14-day incubation period and provided with 10% sucrose and an oviposition cup lined with moist paper towel. After the incubation period, females were individually stored at −80°C. Mosquitoes were dissected to remove the wings for length measurement (iSolution lite; IMT i-solution, Inc.) and bodies and legs were assayed to determine infection and dissemination status, respectively. Here, “infection status” refers to either an infection that is restricted to the midgut (nondisseminated infection) or an infection that has spread beyond the midgut to infect other parts of the mosquito (disseminated infection). Leg assays used here provide a method to determine whether infected mosquitoes are classified as nondisseminated or disseminated infections. Infection and dissemination rates, respectively, were calculated as the percentage of infected bodies and legs from the total number engorged with blood.
The bodies and legs were homogenized in 1 mL of diluent (Leibovitz L-15 media, 10% fetal bovine serum, and 1% penicillin/streptomycin) and 220 μL of mosquito body or legs homogenate were used for RNA extraction using Qiamp® virus Biorobot® 9604 kit according to the manufacturer's protocol (Qiagen). The resulting RNA was reconstituted in 80 μL of TE buffer and stored at −80°C until use. The body and leg samples were analyzed for the presence or absence of SINV RNA using Taqman probe reverse transcriptase–polymerase chain reaction (RT-PCR). PCR amplification was conducted in 25 μL reaction mixture containing 12.5 μL of 2× one-step RT-PCR master mix (Applied Biosystems), 0.65 μL of RNAse inhibitor (Qiagen), 0.25 μL each of 10 μm forward and reverse primers, 0.125 μL of 10 μm Taqman probe, 6.2 μL of nuclease-free water, and 5 μL of template RNA. Primers and probe used for this study targeted the nonstructural protein 1 and were forward primer (5′- CACWCCAAATGACCATGC-3′), reverse primer (5′-KGTGCTCGGAAWACATTC-3′), and probe (5′-FAM-CAGAGCATTTTCGCATCTGGC-BHQ1-3′). RT-PCR was conducted in 7300 real-time PCR system (Applied Biosystems) under the following thermocycling conditions: 50°C for 30 min and 95°C for 5 min, followed by 35 cycles of 95°C for 30 s, 50°C for 30 s, and 72°C for 1 min.
The data were checked for normality and homogeneity of variances, and MANOVA was used to determine the effect of temperature and malathion on infection and dissemination rates. Significant means were separated by Tukey test. Separate analyses of covariance were tested for the effect of size on infection and dissemination rates.
Results
Life history traits and population growth
Survival to adulthood was significantly affected by malathion but not temperature treatment or interaction between malathion and temperature (Table 1). Survival was significantly higher in the absence than in the presence of malathion (Table 2). Male development time to adulthood was significantly longer at 20°C than at 30°C but was not affected by the presence of malathion or interaction (Tables 1 and 2). MANOVA revealed a significant malathion by temperature interaction for female size and development time, with size as the most important variable (Table 3). Female sizes and development decreased with increasing temperature, and at low-temperature larval rearing conditions (20°C), females exposed to malathion were larger than those in the controls (Fig. 1). This effect was not observed at high-temperature (30°C) conditions. An interaction of malathion and temperature also significantly affected λ′ (Table 1). Malathion-exposed mosquitoes had lower λ′ at 30°C than at 20°C (Fig. 1).
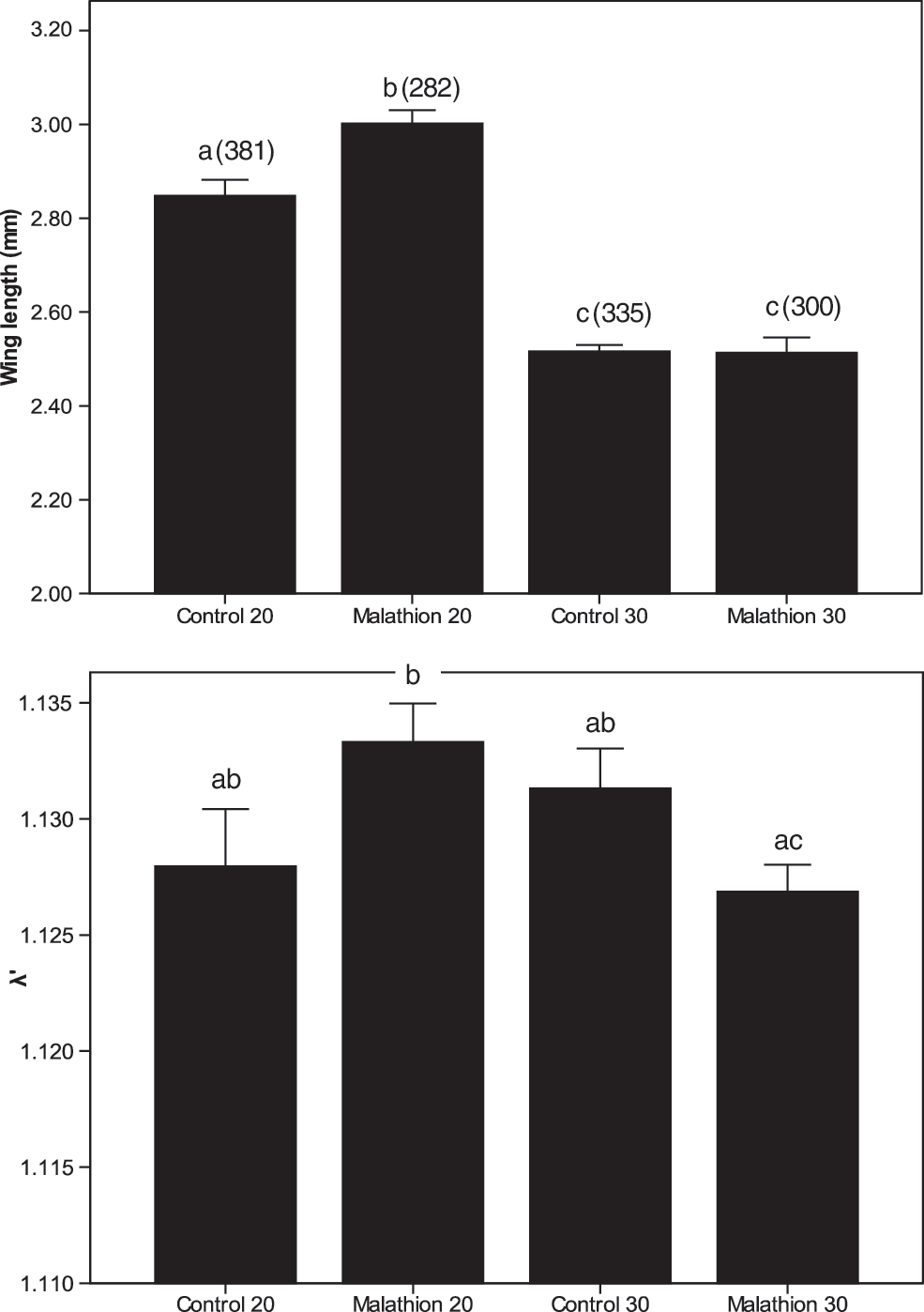
Least squares means±standard error for effect of malathion (0 and 0.05 ppm) and temperature (20°C and 30°C) on female wing length and λ′. Different letters show significant differences between mean values. Numbers above means (in bracket) show the number of mosquitoes tested.
Bold values indicate effects that were statistically significant.
The number of replicates used in calculating means are listed in the column n. Survival data were arcsine transformed to meet the assumption of normality. Untransformed means are presented here for ease of interpretation.
SCC describes the relative contribution of each response variable to significant treatment effects. Negative associations are denoted by (−).
SCC, standardized canonical coefficient.
Vector competence
Infection and dissemination rates were significantly influenced by an interaction between malathion and temperature, with variation in infection largely contributing to the effect (Table 3). The proportion of infected mosquitoes was positively associated with temperature, and at high temperature (30°C), infection rate was significantly higher in malathion-exposed females than in the control treatments (Fig. 2). Similar findings were observed for disseminated infection (Fig. 2). Size as a covariate had no significant effect on infection or dissemination (infection: F=0.04, df=1, 19, p=0.845; dissemination: F=2.14, df=1, 19, p=0.160).

Least squares means±standard error for effect of malathion (0 and 0.05 ppm) and temperature (20°C and 30°C) on percent of females infected and with disseminated infections. Different letters show significant differences between mean values. Numbers above means (in bracket) show the number of mosquitoes tested.
Discussion
The present study used SINV–Ae. aegypti model system to study how temperature and insecticide malathion may alter adult mosquito fitness and interaction with arboviruses. SINV is not naturally transmitted by Ae. aegypti, but it belongs to the same family and genus as chikungunya virus (Togaviridae; Alphavirus), which is naturally transmitted by this species (Thavara et al. 2009). Therefore, this study provides useful insights on how the two environmental variables may affect alphavirus transmission by this species.
Our results demonstrate that temperature can modify the effect of exposure of mosquito larvae to low concentrations of malathion in terms of adult mosquito fitness and overall population performance. Adult female body sizes were negatively associated with temperature, which is consistent with previous studies (Lounibos et al. 2002, Westbrook et al. 2010). Also, the largest adult females were observed in malathion treatments relative to the controls at low temperature. We did not identify the mechanism(s) responsible for these results, but it is possible that reduced survival among immature stages in the presence of malathion released the remaining individuals from competition. Alternatively, malathion may have selectively eliminated small-sized larvae from experimental cohorts, leaving larger-sized females to develop to adulthood. Similar mechanisms were suggested to account for increased body size and fecundity of Ae. aegypti following exposure to spinosad, a naturally derived insecticide (Antonio et al. 2008). Interestingly, there were no differences in mosquito size between malathion and control treatments at 30°C. At higher temperature, rapid larval development may put constraints on resource acquisition and growth, making malathion-mediated size effects less apparent (Korochkina et al. 1997). This may partly account for the higher λ′ observed for malathion-exposed individuals at 20°C compared with the same group at 30°C. These results suggest that low concentrations of toxicants encountered during mosquito larval development have lasting effects on adult traits and are contingent upon the underlying ambient temperature. Therefore, the lethal and sublethal effects of toxicants on mosquitoes may be missed or underestimated in experimental studies conducted at one standard temperature.
Larval rearing conditions had latent effects associated with an interaction between temperature and malathion on SINV virus infection and dissemination rates. Higher temperature increased virus infection and dissemination rates, and malathion-exposed females at this temperature were more competent than those in control treatments. Previous studies have shown that larval rearing temperature (Wada 1974, Mourya et al. 2004, Westbrook et al. 2010) and exposure to insecticides (Noireau and Karch 1983, Seif et al. 1997, Yadav et al. 2005) can affect vector competence for pathogens. Our results further suggest that high temperature and exposure to malathion may increase the ability of mosquitoes to transmit arboviruses. Larger mosquitoes imbibe larger volumes of blood (Klowden and Lea 1978) and, presumably, are more likely to encounter higher doses of virus. In our study, the largest mosquitoes were produced from the cool larval rearing conditions, yet infection and dissemination rates were significantly lower relative to individuals from the warm larval rearing treatment. These results suggest that temperature-induced changes in phenotypic traits, other than body size, were responsible for observed differences in vector competence.
Our study did not address the mechanism by which high temperature and exposure to malathion at larval stage increase vector competence, but we can offer some hypotheses. Higher temperatures, and perhaps exposure to malathion, support rapid development of mosquito larvae and this may reduce their effectiveness to acquire and accumulate nutrient reserves (Korochkina et al. 1997). This may compromise the overall health of the mosquito by causing poor development of some tissues including those related to immunity or reducing the amount of energy allocated to immunity. Previous work with Drosophila melanogaster has shown that higher temperature can indeed impair expression of immune genes (Linder et al. 2008). Further, immunotoxicological studies have shown that pesticides can cause structural and functional alterations to the immune system that may compromise its functions (Blakley et al. 1999a, 1999b). Inhibition of acetylcholine esterase by malathion may also compromise larval feeding behavior, as demonstrated by some insecticides (organophosphates and carbamates) in other aquatic organisms (Pavlov et al. 1992). These factors may jointly or independently account for increased susceptibility to SINV in malathion-exposed individuals at high temperature.
Contamination of surface water with pesticides is very common because of their pervasive use, particularly in agriculture (U.S. Department of Agriculture 1997, El Bakouri et al. 2007). The major ways in which pesticides gain entry into water systems include surface runoff, leaching, spray drift, and/or direct application, particularly when used to control rice pests or aquatic stages of major vectors of medical or veterinary importance such as mosquitoes. Environmental concentrations of these chemicals are often assumed to be nonlethal and unimportant to nontarget organisms. However, our results indicate that exposure of mosquito larvae to low concentrations of insecticide under certain abiotic conditions may cause morphological and physiological changes in resulting adults associated with enhanced vector competence for arboviruses. Our findings may have far reaching implications, given that insecticide use is an integral component of integrated pest management practices. Further, environmental contamination with insecticides may interact with anticipated changes associated with climate change (i.e., increased average temperature) to alter mosquito susceptibility to pathogens. Additional studies are needed to determine whether the observed results translate to other insecticides in determining mosquito susceptibility to pathogens as well as under field conditions.
Footnotes
Acknowledgments
The authors thank M. Dmitrieva, N. Krasavin, A. Montgomery, E. Wachira, and J. Ricci for assistance with daily maintenance of the experiment and R. Lampman for his helpful comments. The authors also thank L.P. Lounibos for providing Ae. aegypti eggs used to start laboratory colonies for this research. This study was supported by the Illinois Waste Tire and Emergency Public Health Funds.
Disclosure Statement
No competing financial interests exist.