
Abstract
The factors involved in the long-term perpetuation of Francisella tularensis tularensis in nature are poorly understood. Martha's Vineyard, Massachusetts, has become a site of sustained transmission of Type A tularemia, with nearly 100 human cases reported from 2000 to 2010. We have identified a stable focus of F. tularensis transmission there, where the annual prevalence in host-seeking Dermacentor variabilis is about 3%, suggesting that this tick perpetuates the agent. However, laboratory studies have shown that infection with F. tularensis has a profound negative effect on dog tick mortality, presenting a paradox: how can a vector perpetuate an agent that negatively affects its fitness? It may be that experimental infection does not mimic that of natural transmission. Accordingly, we examined the effects that F. tularensis has on the longevity of field-derived ticks. Of 63 PCR-positive ticks collected in early summer, 89% were dead by December compared to 48% of 214 uninfected ticks collected at the same time and site. However, the quantum of F. tularensis DNA within each tick was not correlated with increased mortality. Instead, ticks with an uncommon genotype were more likely to die early than those with the common genotype. We conclude that the interaction between F. tularensis and its vector is complex and certain bacterial genotypes appear to be better adapted to their arthropod host.
Introduction
Martha's Vineyard, Massachusetts, has sustained a tularemia epizootic of unusual duration, starting in 2000 with as many as 100 human cases cumulatively reported through 2010, ranging from 3 to 16 cases a year (Matyas et al. 2007). Indeed, it would be more appropriate to characterize this 237 km2 island as tularemia endemic. We have previously determined that American dog ticks (Dermacentor variabilis) are commonly infected and that stable microfoci of transmission exist (Goethert et al. 2004, Goethert and Telford 2009). Although tularemia has long been classified as an infection of natural nidality (Pavlovsky 1966), to our knowledge our findings are the only demonstration that a discrete site may be sampled year after year and always yield evidence of active F. tularensis tularensis transmission. Such stability suggests that there is efficient vertical and/or horizontal transmission by ticks, thereby sustaining the microfocus. However, vertebrate hosts are either scarce (rabbits, voles, and white-footed mice), perhaps due to tularemia mortality, or are immune [skunks and raccoons (Berrada et al. 2006)]. An environmental reservoir (water or soil) has not been found (Berrada and Telford 2010). The biological basis for the persistence of a natural focus remains undescribed.
It may be that ticks serve as a long-term reservoir in the Martha's Vineyard natural foci and that regulation of enzootic tularemia depends on factors impacting the ticks. A seminal study of tick competence for F. tularensis (Philip and Jellison 1934) as well as a recent study (Reese et al. 2010) demonstrated that infected ticks die, similar to the effects on fitness seen for Rickettsia rickettsii in wood ticks (Niebylski et al. 1999). The degree to which ticks serve as long-term reservoirs would depend on the prevalence and intensity of a negative fitness effect. The fact that we have found infected ticks in our study sites every year for nearly a decade poses a paradox with the experimental data. It may be, however, that experimental transmission does not entirely simulate infection acquired under natural circumstances. We incidentally observed unusually great rates of mortality of field-collected ticks that had been initially screened as PCR positive for F. tularensis. Accordingly, we compared the longevity of naturally infected (as determined by PCR of nondestructively sampled hemolymph) ticks with those that did not contain evidence of F. tularensis DNA. In addition, we determined whether the bacterial burden within ticks was associated with mortality. Finally, because there is a great diversity of F. tularensis tularensis variable number tandem repeat (VNTR) genotypes present on Martha's Vineyard (Goethert et al. 2004), we tested the hypothesis that certain bacterial genotypes were associated with mortality.
Materials and Methods
Tick collection and PCR
Host-seeking dog ticks were collected by dragging vegetation on our field site near Squibnocket on Martha's Vineyard, from April to June 2006. This site has previously been described (Goethert and Telford 2009). Ticks were tested for evidence of F. tularensis DNA by the hemolymph test. The distal tip of one of the forelegs was clipped with flamed surgical scissors, and a drop of hemolymph was collected into a sterile microcentrifuge tube, which was assigned a unique pool identification number. Hemolymphs from six ticks were pooled, centrifuged, and boiled at 100°C for 10 minutes. An aliquot of the boiled suspension was used directly for PCR targeting the fopA gene, as described previously (Goethert et al. 2004).
Longevity study
The ticks were analyzed by pooling hemolymphs from six ticks, which were held individually within separate new 1.5 mL microfuge tubes (Eppendorf ) after hemolymph was collected; each pool was then given a unique identifying number. When a pool of hemolymph was found positive, all the ticks from that pool were then enrolled in the study without further knowledge of infection status. After each timepoint, dead ticks were then extracted for DNA and assayed for evidence of F. tularensis infection by PCR, as were any remaining at the end of the observation period. The sample size for the study was not chosen a priori: we included all of the hemolymph-positive ticks from one specific site [our “natural microfocus” (Goethert and Telford 2009)]. The comparison group comprised the uninfected ticks of the pools of six. Thus, we used 46 pools of six, with 1 pool comprising seven ticks. Although many more ticks were collected during the season from the site, we only included those that were analyzed at the same time from the same collections so that they all experienced the same conditions at the same time.
Tubes were held vertically in a standard 80-well polystyrene microcentrifuge rack, which in turn was held within an acrylic dessicator chamber containing a beaker of distilled water. The dessicator chamber was held in a BOD cabinet (Thermo Model 815) at a constant 15°C with no light; the relative humidity within the dessicator chamber was 95%. Ticks were checked for viability in September and again December. Dead ticks were removed and tested individually for evidence of infection. Whole ticks were homogenized in 50 μL brain heart infusion broth using a flame-sealed pipette tip and a 12 μL aliquot was extracted for DNA using Isoquick (Orca Research, Bothell, WA) according to the manufacturer's recommendations. The template was tested for evidence of F. tularensis DNA by PCR targeting the fopA gene (Goethert et al. 2004). Any tick that remained alive at the end of the observation period was tested for evidence of infection by the hemolymph test and PCR.
Quantitative real-time PCR
Real-time PCR targeting the tul4 gene was then used to determine the number of genome equivalents of F. tularensis tularensis in individual tick samples, as previously described (Goethert and Telford 2010). The 28S ribosomal RNA gene for D. variabilis was used as an internal standard, and a 200bp synthetic oligo was synthesized for use as a positive control. The results are expressed as genome equivalents per tick, as previously described. Only 46 of the 63 ticks that were initially PCR positive were analyzed for quantum of infection; we extracted DNA only from ticks that were considered freshly dead to ensure that estimates of bacterial burden were not confounded by issues of DNA degradation.
Genotyping of F. tularensis tularensis in Dermacentor variabilis
The F. tularensis tularensis in PCR-positive ticks were genotyped using VNTR analysis as described previously (Goethert et al. 2004, Johansson et al. 2004). Briefly, two loci, Ft-M3 and Ft-M10, were amplified using a fluorescently labeled primer and a high-fidelity Taq polymerase (Picomaxx, Stratagene). Amplicons were mixed with 500bp ROX-labeled marker (Applied Biosystems) and sent to the University of Maine Sequencing facility for accurate sizing using GeneMapper analysis.
Statistical analysis
This study was not an experiment that had been designed a priori, but was initiated as an observation of ticks that were stored alive pending final analyses. Groups of ticks collected from the field during the spring of 2006 were examined at two time points and those that were dead at those time points were removed for further analysis. Thus, the design could be considered either a case–control (infected or not) study at each of the two time points or as a longitudinal cohort study (cumulative status of the infected and uninfected groups). The data are presented as contingency tables and χ 2 tests (Yates correction) performed. The analyses were considered significant a priori if p<0.05.
Results
Two hundred seventy-seven Dermacentor variabilis were included in the study. By September, 44% of the 63 PCR-positive ticks had died, whereas only 24% of the 214 uninfected ticks died (p<0.05, Table 1). At the December observation point, 80% of the remaining 35 PCR-positive ticks were dead as opposed to 32% of the remaining 163 uninfected ticks (p<0.001, Table 2). Cumulatively, by the end of the observations, 89% of the 63 PCR-positive ticks had died, whereas only 48% of the 214 uninfected ticks had died (p<0.001, Table 3). We conclude that natural infection with F. tularensis tularensis contributes to the mortality of D. variabilis, at least when the ticks are held in a laboratory setting.
χ 2=9.2, df=1, p<0.05.
χ 2=25.7, df=1, p<0.001.
χ 2=31.4, df=1, p<0.001.
To determine whether ticks that died in September were more likely to have a greater degree of infection (bacterial burden) than those that died in December, we quantitated F. tularensis DNA in each tick by quantitative real-time PCR, and in particular, compared the amounts of specific DNA in ticks that had died by September 2006 (n=20) with those dead by December (n=26). No significant difference in the amount of F. tularensis DNA (and thus potential bacterial burden) was detected (p=0.28 Wilcoxon two sample test) between the two groups. Those ticks that remained alive at the end of the observations did not differ with respect to bacterial burden from those that were dead (Fig. 1).
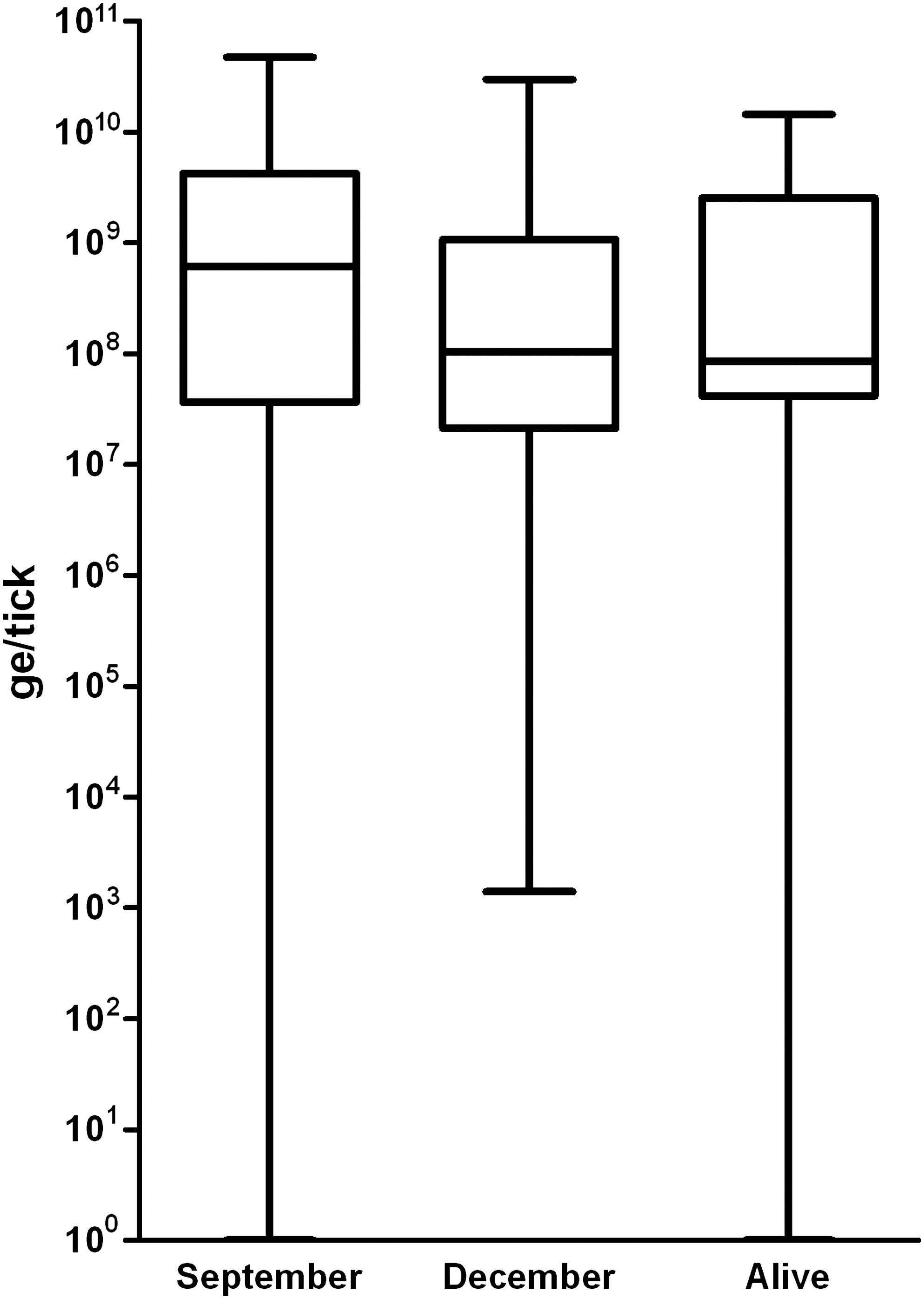
Boxplots for F. tularensis genome equivalents in ticks that died in September compared to those that died in December as well as to those infected ticks that remained alive at the end of the observation period.
Finally, we typed the F. t. tularensis using two VNTR markers described previously. Our previous work with this population of ticks has identified 13 different genotypes using these two markers. Most ticks (69%) contain a single genotype, 10 7. The other 12 genotypes comprise the remaining 31% of infected ticks (Fig. 2). The proportion of ticks containing 10 7 for our study is the same as that for our previous sampling, suggesting that our study was representative of what is usually found in our study site. For the purposes of this study, we divided the F. t. tularensis strains into two groups: the common genotype, 10 7, and the uncommon genotypes. We then determined whether ticks from either group were more likely to die early, that is, in September as opposed to December. At the September sampling, 61% of those ticks with uncommon VNTR bacteria had died compared to 31% of those with the common 10 7 VNTR type (p<0.05, Table 4). By the December timepoint, there was no apparent difference in mortality between ticks containing either VNTR type (Table 5). There was no difference in mortality between uninfected ticks and those containing the common 10 7 VNTR (p=0.452, Table 6), but there was a clear difference in mortality between those containing uncommon VNTRs and uninfected ticks (p<0.001, Table 7), suggesting that the uncommon VNTRs greatly contributed to the overall differential mortality. We conclude that the uncommon F. tularensis tularensis genotypes are associated with greater mortality of infected ticks and that genotype 10 7 may represent a lineage that has modulated its effects on tick fitness.
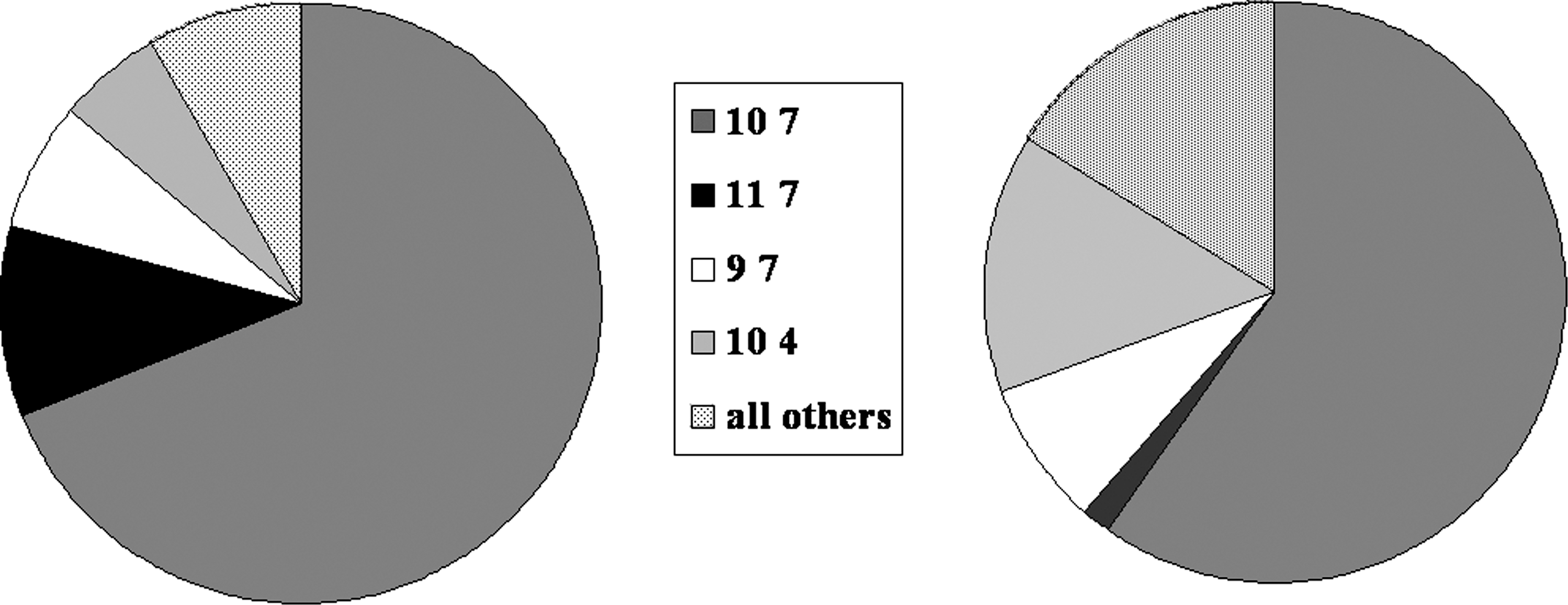
The distribution of VNTR genotypes detected in dog ticks (2002–2006) from our study site (left panel) compared with that for ticks in this study (right panel).
χ 2=4.3, df=1, p<0.05.
χ 2=0, df=1, p=1.00.
χ 2=0.57, df=1, p=0.452.
χ 2=14.9, df=1, p<0.001.
Discussion
Deleterious effects of infection on vectors, such as increased mortality or diminished fecundity, have been demonstrated for diverse pathogens such as arboviruses or malarial parasites (Cooper et al. 2000, Moncayo et al. 2000, Watt and Walker 2000, Ochanda and Young 2003, Fernandez et al. 2003, Montoya-Lerma et al. 2003, Ross and Levin 2004, Styer et al. 2007). The agent of Rocky Mountain spotted fever is a particularly relevant example because it too is maintained by Dermacentor spp.ticks. A large proportion of experimentally infected D. andersoni died before molting (Niebylski et al. 1999) and infected females produced fewer progeny, if at all. Interestingly, not all R. rickettsii transmission studies have documented such negative fitness effects (Telford 2009). The biological basis for some experiments documenting negative fitness effects of infection as opposed to others that do not remains unexplored.
Mortality of ticks due to infection by F. tularensis is not a novel finding (Philip and Jellison 1934, Petrov 1960). Philip and Jellison (1934) noted that there was mortality of D. variabilis in their experiments examining transstadial and transovarial transmission (TOT) but failed to quantify the amount. In contrast, infected D. andersoni nymphs were able to maintain and transmit infection after 6–8 months (Bell 1945) and unfed infected adults were able to live 5 months, but mortality was not recorded (Parker et al. 1924). Hopla (1953), using Amblyomma americanum, did not mention whether infected ticks tended to die. It is thus difficult to generalize from these previously published reports because they used different species and stage of ticks; small sample sizes or limited quantitation of the fitness effect further restricting generalization. In addition, most of the previous experiments were conducted before the formal recognition of at least two F. tularensis subspecies (F.t. tularensis and F.t. holarctica) on the basis of differences in virulence and epidemiology. It is not clear which subspecies were used in each experiment.
A recent report (Reese et al. 2010) greatly clarifies these issues and corroborates the negative fitness effects of previous transmission studies. Using isolates representing F. tularensis tularensis clades A1b (a strain isolated from the sole Martha's Vineyard pneumonic tularemia fatality and in fact comprising VNTR 10 7) and A2 as well as F. tularensis holarctica, the dynamics of transstadial transmission were examined using an Oklahoma strain of D. variabilis. There was a difference in overall survival of nymphs infected by clade A1b compared to uninfected ticks, 59% of 368 uninfected versus 35% of 912 infected surviving at day 65. In addition, infected nymphs took more than twice as long to feed as uninfected and fewer than half fed to repletion. These are profound negative fitness effects.
The conditions in which we held the ticks over the course of the experiment were not intended to replicate those to which ticks are naturally exposed. However, all ticks in this experiment were handled in the same manner, being collected from the same site at the same time, initially processed at the same time and held in the same environmental chamber. The average temperature recorded every 12 minutes by a datalogger placed at ground level in our study site, for 2007, was 16.8°C from March 15–October 15. Thus, 15°C is representative of the average temperature to which naturally infected ticks are exposed. Whether photoperiod may influence the interaction of the pathogen with its tick host is not clear, although photoperiod clearly triggers the entry and exit of a behavioral diapause for D. variabilis (Sonenshine 1991). Additional experimental studies in which natural conditions are better simulated in the laboratory are warranted inasmuch as placing experimentally infected ticks into the field within cages would not be possible given the Select Agent status of F. tularensis.
The factors that limit F. tularensis tularensis transmission in nature remain poorly explored. Vector longevity has significant influence in quantitative models of vectorial capacity (Hartemink et al. 2008) and thus greatly impacts the basic reproduction number of the pathogen transmitted by a vector. In addition, a negative fitness effect of infection would argue against perpetuation by TOT; in general, agents that cause deleterious fitness effects for their vector are not maintained vertically and instead rely on a horizontal transmission cycle for amplification (Lambrechts and Scott 2009). The extent to which TOT contributes to the perpetuation of F. tularensis remains unresolved. If TOT is rare, adult ticks only indirectly contribute (tick reproduction only) to the basic reproduction number of F. tularensis tularensis because this stage rarely feeds on hosts upon which subadult ticks feed (at least, in our study site) and are thus unlikely to contribute to transstadial transmission (either systemically or by cofeeding). However, adult dog ticks are of zoonotic importance because they attach to humans, whereas subadult dog ticks do not (Smith et al. 1946).
As reported by Reese et al. (2010), mortality was not associated with the bacterial load within the tick. We found no significant association between the quantum of infection and the proportion of ticks that survived. One of the basic tenets of infection biology is that pathology may be a function of pathogen burden. Why “disease,” as expressed by decreased survival of infected ticks, appears to be causally unrelated to bacterial burden remains unclear and indeed suggests that there is a complex mechanistic basis for the phenomenon. Even some of the ticks with the lowest burden of F. tularensis died early.
Instead of bacterial burden, we found that bacterial genotype was associated with the mortality of PCR-positive ticks. Ticks with the uncommon genotypes disproportionately contributed to the mortality of infected ticks at the September observations. The molting success of deer ticks may be altered by strains of Anaplasma phagocytophilum (Ross and Levin 2004), and different Plasmodium sp. strains have varying effects on the mortality of their mosquito vector (Ferguson and Read 2002). Pronounced strain differences in mortality of the vector would create an imbalance in the basic reproductive number for the each strain. This alone could be responsible for the dominance of the 10 7 genotype over the uncommon F. tularensis types in our studies on Martha's Vineyard (Goethert et al. 2004, Goethert and Telford 2009, Goethert et al. 2009). Interestingly, environmental factors (e.g., availability of glucose) had a larger effect than mosquito genotype on the ability of mosquitoes to resist infection with malaria parasites (Lambrechts et al 2006). Although ticks do not feed on sugar in between blood meals, it may be that other environmental factors, such as relative humidity, temperature, or insolation, will have a mitigating effect on the survival of infected ticks. Mortality was reduced, for example, when Theileria parva infected ticks were held in quasi-natural conditions as opposed to standard laboratory conditions (Ochanda and Young 2003).
The mode of perpetuation of most tick-borne pathogens remains incompletely understood. Of particular interest is whether the tick or a vertebrate host serves as the reservoir or critical factor that ensures that the pathogen's basic reproduction number exceeds unity. At one end of a possible continuum of scenarios, an agent may be stably inherited and passed through each tick life stage with no need for transmission to a vertebrate, as is the case for endosymbiotic rickettsiae. On the other end of the continuum are agents such as North American Lyme disease spirochetes, which are transmitted in the nymphal stage to susceptible hosts, which then serve to infect a new cohort of larvae. Intuitively, the most adaptive strategy would be a combination of inheritance (transovarial or vertical transmission) with the requirement for a reservoir or amplifying host (horizontal transmission). Both modes operating simultaneously would ensure perpetuation: in the event that reservoir hosts are scarce, ticks vertically maintain the agent and if infected tick lineages are killed as a result of catastrophe, a single infected tick might generate multiple new lineages of infected ticks via an amplification host. Even so, considerable debate exists regarding the relative roles of vertical and horizontal components for the perpetuation of many tick-borne pathogens. Besides the basic interest in understanding the evolutionary ecology of tick-borne pathogens, such issues influence the likelihood of success of intervention strategies such as reservoir targeted vaccines or tick control.
Footnotes
Acknowledgments
Our laboratory is funded by a grant from the National Institutes of Health (R01 AI 064218). We thank the Martha's Vineyard residents and Vineyard Open Land Foundation for allowing access to their properties, and John Varkonda of the Massachusetts Department of Conservation and Recreation for logistical support.
Disclosure Statement
The authors have no conflicts of interest to disclose.