
Abstract
Hantaviruses are distributed throughout the United States and are the etiologic agents for hantavirus pulmonary syndrome and hemorrhagic fever with renal syndrome. Hantavirus genotypes and epidemiologic patterns vary spatially across the United States. While several longitudinal studies have been performed in the western United States, little is known about the virus in the eastern United States. We undertook a longitudinal study of hantaviruses in the primary rodent reservoir host in central Pennsylvania, Peromyscus leucopus. Prevalence of hantavirus antibodies varied both by year and site, but was not correlated with host abundance. Males were significantly more likely to have antibodies to a hantavirus than females, and both antibody sero-conversion and antibody prevalence increased with mass class (indicator for age). Our findings suggest that one or more hantaviruses are present and circulating among P. leucopus of central Pennsylvania, and understanding the dynamics in this region could help prevent zoonotic transmission to humans. Our aim was to describe the differences in epizootiology of hantavirus infection in rodents from various geographical locations to enable improved analysis of the risk of rodent-to-human transmission and obtain insights that may indicate improved means of disease intervention.
Introduction
Transmission of hantaviruses to humans occurs most frequently through the inhalation of aerosolized virus from dried feces, urine, and nest sites of rodent hosts. Hence, the epidemiology of hantaviruses in North America is closely linked to the ecology and infection of rodent reservoirs. By understanding the risk factors associated with hantavirus infection in rodent hosts, and the temporal and spatial variation in transmission, we can improve predictions of relative risk to humans. Several factors have been associated with the presence of antibody to hantaviruses in rodent hosts, including host gender and mass (indicator for age) (Mills et al. 1997, Calderon et al. 1999, Cantoni et al. 2001, Douglass et al. 2001, Escutenaire et al. 2002, Calisher et al. 2007). Further, an association between the presence of wounds and antibody prevalence has been demonstrated (Douglass et al. 2001, Glass et al. 2002, Calisher et al. 2007), suggesting that transmission between rodents may occur via aggressive encounters. As such, we would expect transmission to increase with host population density (Mills et al. 1999, Kuenzi et al. 2007, Luis et al. 2010); however, empirical evidence for this type of association has been equivocal. A central hypothesis from arid regions is that rainfall increases vegetation and food for the mice, altering the structure of the predator community, thus increasing the likelihood of zoonotic transmission (Glass et al. 2002). In 1993 a significant outbreak in the Four Corners region of the western United States was linked with a rapid increase in rodent population density associated with increased precipitation (Parmenter et al. 1993, Mills et al. 1999). This type of trophic cascade is unlikely to happen in northern hardwood forest of Pennsylvania, where the mice exhibit regular oscillations in abundance (Wolff 1996).
Human cases of HPS have occurred in the northeastern United States, with cases recorded from Maine, New York, Pennsylvania, Maryland, Virginia, and West Virginia (Rhodes et al. 2000). Clearly, hantaviruses persist within the region and probably within Peromyscus populations; indeed, this was confirmed when a study in Pennsylvania recorded 17% of captured Peromyscus spp. had antibodies to a hantavirus (Mills et al. 1998). Given that the public health significance and epidemiology of this infection is likely to vary between ecological habitats, we undertook a preliminary study to record spatiotemporal variation in antibody prevalence in Peromyscus leucopus populations from central Pennsylvania. Specifically, we asked the following: Does antibody prevalence in the host population vary among locations and over time? Is antibody prevalence associated with host population abundance within and among sites? Was antibody prevalence associated with host factors such as gender, age, and wounding status? Ultimately, these data can be used to develop models for predicting disease risk and spillover events into human populations.
Materials and Methods
Verification of Peromyscus species
The geographical range of two morphologically similar species, P. leucopus and P. maniculatus, are known to overlap in Pennsylvania. As such, we performed DNA sequencing on a subsample of the mice (n=12) to verify the species of Peromyscus in our study sites. See Ivanova et al. (2007) for detailed methods. Briefly, we collected tail snips from mice trapped in the field and preserved the tissue samples in dimethyl sulfoxide. DNA was extracted from the tissue samples and amplified with a COI-2 (cytochrome c oxidase subunit 1) primer cocktail (Ivanova et al. 2007). PCR products were sequenced using the BigDye Terminator version 3.1 Cycle Sequencing Kit (Applied Biosystems, Inc.) on an ABI 3730 capillary sequencer. Sequences were viewed on Sequencing Analysis Software version 5.1.1 (Applied Biosystems). The sequences were submitted onto BOLD-IDS (Bold Systems version 2.5) for species identification.
Field collection
Peromyscus mice were trapped biweekly from May to September in 2005, 2006, and 2007 from three mix-hardwood forest sites located 20 km south of State College, Pennsylvania, referred to as Yellow Hickory, Broken Arrow, and Rothrock. Each site had three grids separated by at least 200 meters, for a total of nine grids. Each grid consisted of 64 multi-capture live traps (Ugglan, Graham) located at 10 m intervals in an 8×8 configuration. Traps were set on two consecutive nights; if mice were recaptured on the second night, only trap location was recorded before release. On first capture, mice were tagged with a passive integrated transponder (Trovan™; EIDAP) for identification of individuals. Trap location, PIT tag number, body length, body mass, sex, and presence of wounding (torn ears, cuts, and visible scars) were recorded. Once an animal had been processed, it was returned to the trap location and released. Field collection was conducted with the approval of the Pennsylvania State University Animal Care Committee (IACUC #16061).
Trap data were used to estimate minimum number known alive (MNA) each month in the mouse population. This index was calculated by taking the total number of individual mice captured during each 2-day trapping session and adding to that the number captured on at least one previous and one subsequent session, but not during the month of interest (Krebs 1966). Blood samples were collected from live mice by retro-orbital bleeds and stored on ice until centrifugation. After separation, red blood cells and serum were stored separately at −20°C. Serum samples were sent to the Centers for Disease Control and Prevention to be tested for antibody reactive with Sin Nombre virus (SNV) recombinant nucleocapsid protein by enzyme-linked immunosorbent assay (see Mills et al. 1999 for details). Note that antibody test did not differentiate between different strains of the SNV virus.
Statistical analyses
Data were analyzed using the statistical package R (
MNA was analyzed with a Poisson error distribution to determine whether mouse abundance varied between months, years, and sites. An a posteriori Tukey test was performed to delineate specific differences where an overall effect was significant. To test the relationship between host abundance and hantavirus antibody prevalence, a separate analysis on antibody prevalence was performed: the response variable was proportion of mice antibody-positive, and the explanatory variables were MNA, site, month, year, and their interactions.
In analyzing the relationship between age and infection, individuals were grouped into mass classes following the procedures used by Vandegrift and Hudson (2009): class 1<16 g (juvenile status); class 2=16–18 g; class 3=18.1–20 g; class 4=21.1–23 g; class 5>23 g. If an animal appeared more than once within a single mass class, only the first antibody-positive sample was included in the analysis; if both were negative, then a single negative entry was used (mice that became positive for the presence of hantavirus-specific antibodies rarely reverted back to a negative status). To test whether the age–prevalence curve fit a linear or polynomial function, we analyzed the following general linear mode: antibody prevalence ∼ mass + mass2 + sex, with binomial errors.
To assess the relationship between aggression and host characteristics, wounding (proportion of animals with wounds or scars) was analyzed with mass, mass2, and sex entered into the model as explanatory factors. In a separate analysis, we tested the effect of host mass class and wounding status on incidence (proportion of sero-conversions per mass class). As described above, a mouse was only entered once per mass class. In general, the minimal adequate model was selected using backward-stepwise deletion, retaining a variable if the drop in deviance was significant (p<0.05 based on chi-square statistics).
Results
All 12 of the tail snip samples matched the species P. leucopus with a probability of placement between 99.1% and 99.8%. Hence, it is highly likely that the dominant species in our study sites is P. leucopus, which is consistent with the morphological determination in the field.
The total number of trap-nights for 2005, 2006, and 2007 were 1920, 2560, and 1792, respectively. In most cases only 1 mouse was captured per trap. In addition to P. leucopus, other nontarget rodents were also caught in the traps, including Tamias striatus, Clethrionomys gapperi, Microtus pinetorum, and Blarina brevicauda; blood samples were not taken from these animals. We collected and examined a total of 2142 serum samples from P. leucopus: 741 samples from 379 individuals in 2005, 941 samples from 495 individuals in 2006, and 460 samples from 282 individual mice in 2007. The average prevalence of detectable hantavirus antibody in Peromyscus was 5% (range: 0%–16%), and varied by year and site (Fig. 1).
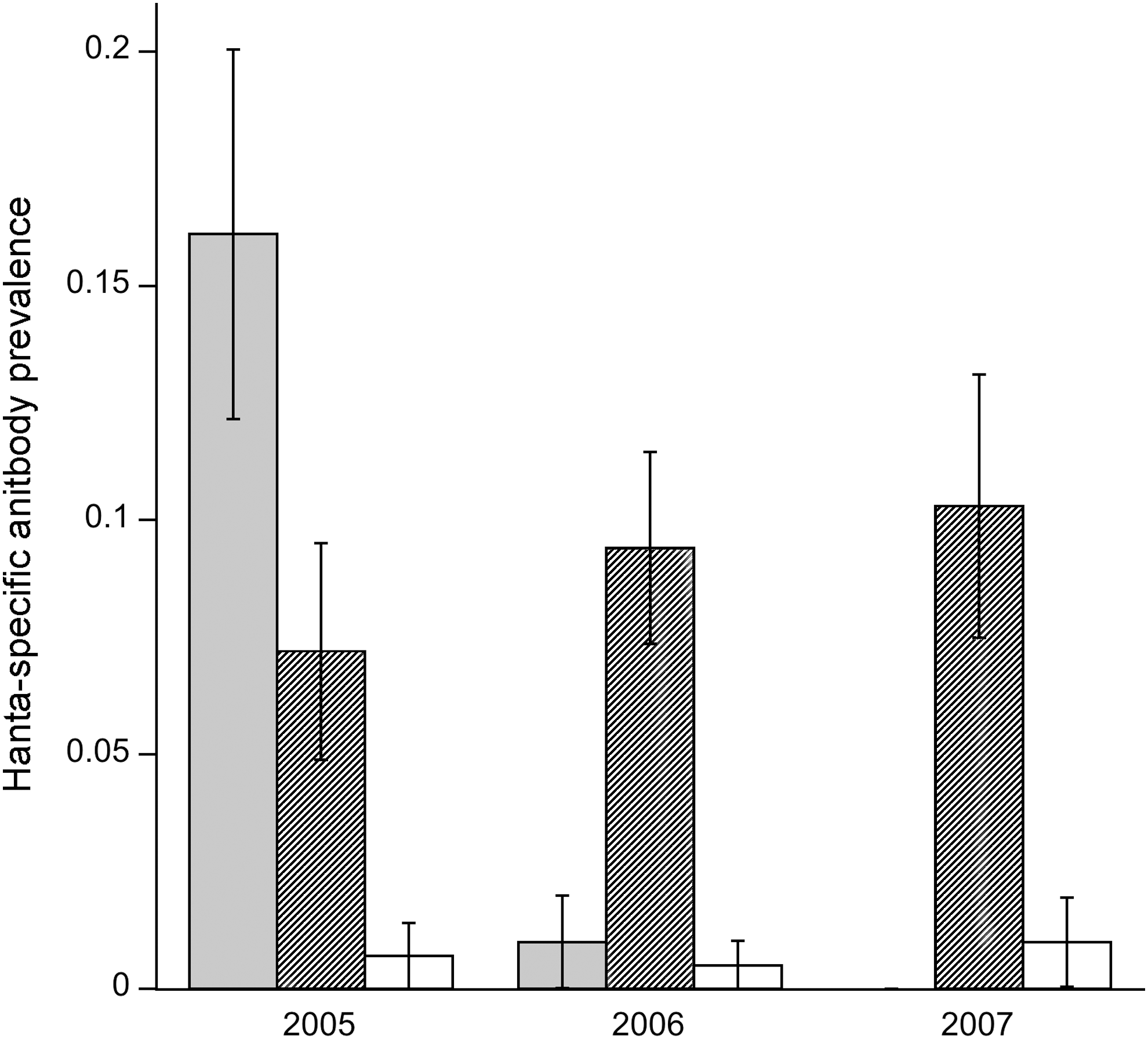
Prevalence of hantavirus antibodies across three study sites over 3 years. The three sites examined were Broken Arrow (gray bars), Yellow Hickory (hatched bars), and Rothrock (white bars).
Overall, year alone was not a statistically significant factor (deviance [=sum of squares]=1.49, p=0.22), but site (deviance=39.3, p<0.001), sex (deviance=10.5, p<0.001), and the interaction between year and site (deviance=23.3, p<0.001) were significant factors; month was not significant and was deleted from the model. The overall sex ratio (male:female) was 1.27 and was not statistically different by site (deviance=3.94, p=0.14) or year (deviance=1.19, p=0.13). Overall, Rothrock had the lowest prevalence for all years (pooled across all months) with <1% of mice found positive for hantavirus-specific antibodies. Broken Arrow had the highest prevalence, with 16.1% of mice expressing hantavirus-specific antibodies in 2005; however, in subsequent years antibody prevalence dropped below 1%. In contrast, antibody prevalence remained fairly constant across all 3 years in Yellow Hickory with a range of 7.2%–10.3% (Fig. 1).
Mouse abundance, estimated as MNA, varied by site (deviance=61.7, p<0.001), but not year (deviance=1.85, p=0.17); however, the site–year interaction was significant (deviance=23.7, p<0.001); month was not significant and was deleted from the minimal model. An a posteriori Tukey test by site and year (pooled across all months) showed that in the first year, mouse abundance was significantly higher in Rothrock (p<0.001) and Yellow Hickory (p<0.001) than in Broken Arrow. However, there was no significant difference in mouse abundance between sites in the following year. In the third year of the study, abundance reached its highest level in Yellow Hickory, and was significantly higher than either of the two sites (p=0.01, p=0.01). The proportion of mice antibody positive for a hantavirus in a given site and year was not associated with MNA (deviance=0.03, p=0.68). Even when the site with near-zero antibody prevalence (Rothrock) was excluded from the analysis, mouse abundance still did not predict antibody prevalence (deviance=0.05, p=0.81).
Prevalence of hantavirus-specific antibodies increased linearly with mass class, which served as a proxy for age (Coefficient=0.87, standard error [SE]=0.25, p<0.001) (Fig. 2). Antibody prevalence was significantly higher (deviance=10.5, p<0.001) in male mice (7.1%±1.0% s.e.) than in female mice (2.7%±0.7% s.e.). Other host factors such as wounding also increased with host mass class: the data fit a second-order polynomial function (linear coefficient: 1.22, SE=0.25, p<0.001; quadratic coefficient: −0.14, SE=0.04, p<0.001) (Fig. 3). Wounding, however, was not correlated with host sex (linear coefficient: 0.12, SE=0.14, p=0.36). Although incidence (proportion of mice found positive for the first time) increased with mass class (deviance=17.04, p<0.001), it was not directly correlated with wounding (deviance=0.15, p=0.70) (Fig. 4). Wounds appeared to have healed in 6% of the mice (out the 426 individuals with mutliple captures), as they were not detected in subsequent trap sessions.
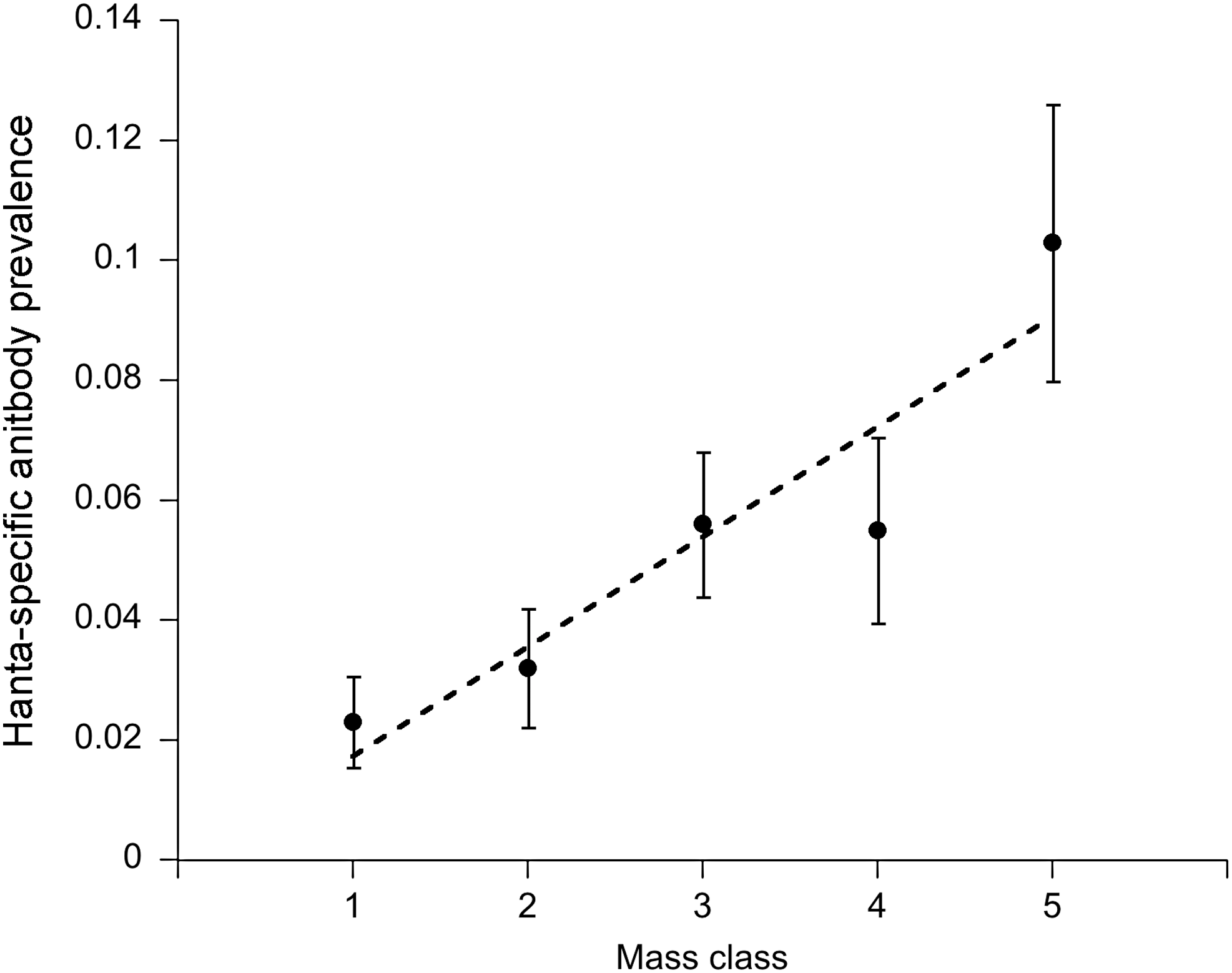
Age–prevalence curve for hantavirus infection in Peromyscus leucopus. Mice were assigned to age classes according to mass: 1 (<16 g), 2 (16–18.9 g), 3 (19–21 g), 4 (21.1–23 g), and 5 (>23 g). The dashed line is a fitted linear regression for hantavirus-specific antibody prevalence in relation to host age.
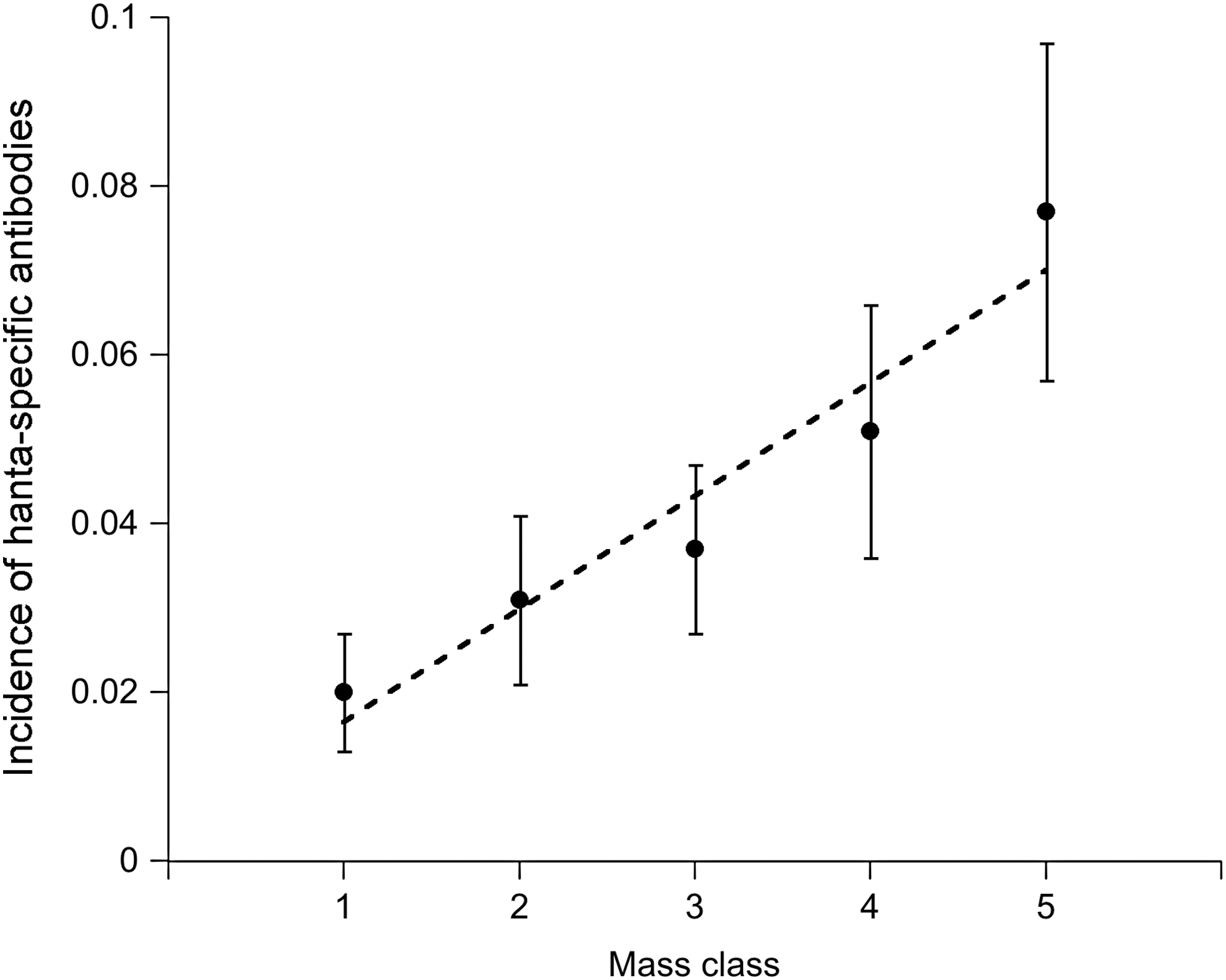
Relationship between mass class (indicator for age) and incidence (proportion of first time antibody-positive mice) of hantavirus in P. leucopus. The dashed line is a fitted linear regression for hantavirus-specific antibody incidence.
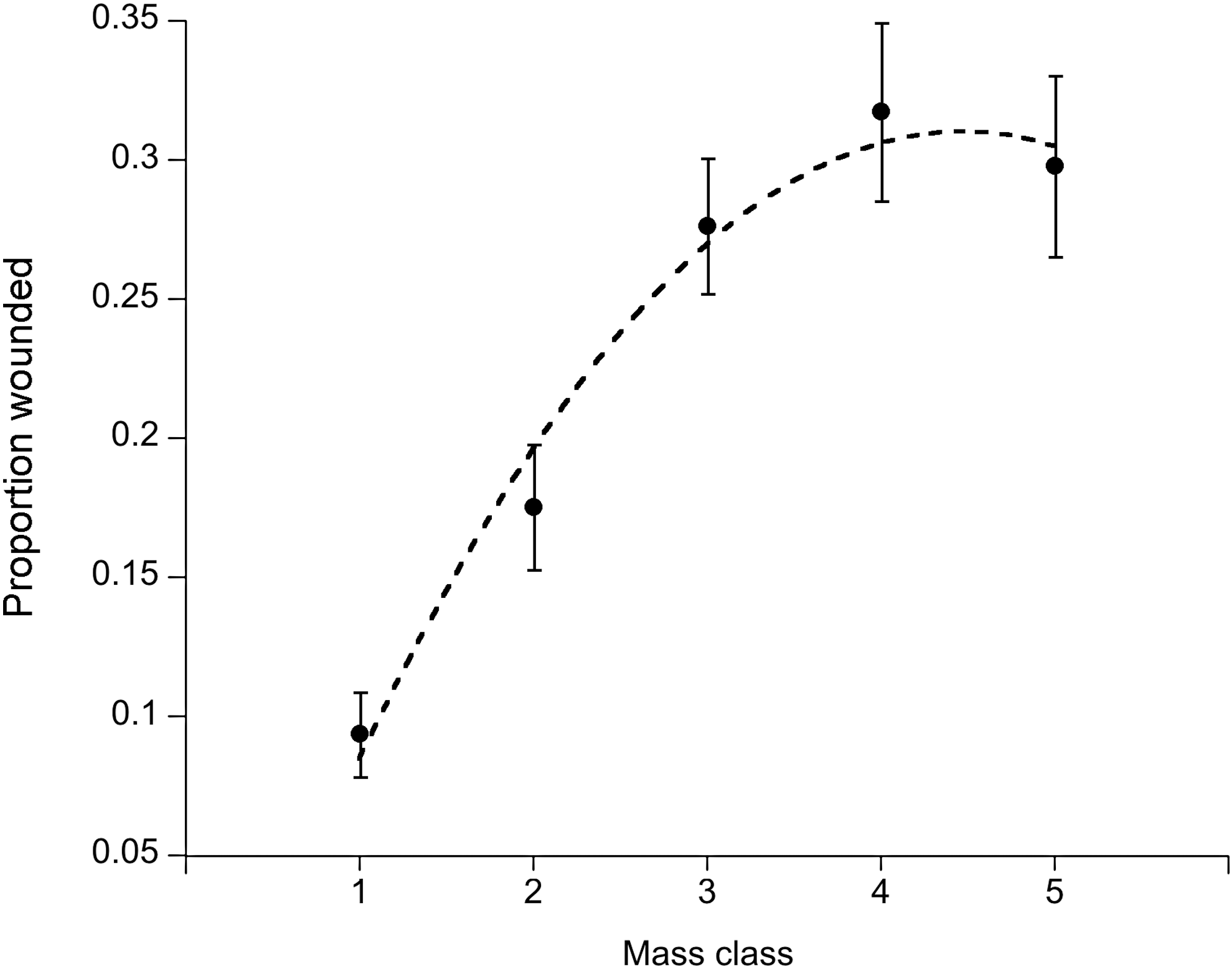
The relationship between wounding status and host mass class (indicator for age). Mice were grouped into mass classes representing age groups. The dashed line represents a best fit second-order polynomial fit to the data.
Discussion
The overall aim of this study was to understand the ecological factors associated with hantavirus infection in Peromyscus leucopus, an important reservoir host. In this study, we examined demographic factors, including host sex, age, and wounding, associated with risk of infection. Prevalence of hantavirus-specific antibodies was positively associated with host mass (a proxy for age) and sex, such that older males were more likely to be antibody positive. We also detected spatial and temporal variation in antibody prevalence in populations of central Pennsylvania over a 3-year period. The average antibody prevalence (5%) was lower than values reported in other studies 7% to 17% (Boone et al. 1998, Mills et al. 1998, Douglass et al. 2001, Kuenzi et al. 2007). Although both antibody prevalence and mouse abundance varied among sites and years, the MNA did not serve as a good predictor of hantavirus antibody prevalence in our study.
Several studies have also failed to demonstrate a simultaneous positive association between hantavirus antibody prevalence and Peromyscus population density (Mills et al. 1997, Douglass et al. 2001); still, Calisher et al. (2007) reported a positive correlation between infection and density. Whether or not an association was found may be in part due to the duration of the study as the longer study found a positive correlation with density, which might be more apparent with additional time points, especially if the correlation with density is delayed as has been observed previously (Luis et al. 2010). In our study, examination of a larger area and extending the time period of the study could help reveal a relationship between density and prevalence. In addition to environmental factors such as precipitation and temperature, differences in the microhabitat use may also influence transmission of hantaviruses. Goodin et al. (2009) found a significant difference in the microhabitat occupied by antibody-positive compared to antibody-negative rodents, suggesting that the risk of transmission and maintenance of hantaviruses is greater in microhabitats with dense canopy cover (Goodin et al. 2009). Therefore, future studies of hantaviruses in Pennsylvania should involve longer-term surveys that measure climatic factors and microhabitat associations.
We did not observe any significant seasonal or monthly variation in antibody prevalence. Although the prevalence of hantavirus antibodies in different grids fluctuated on a monthly basis, these fluctuations in grids did not form any distinguishable pattern and were not consistent within sites from year to year. In contrast, previous studies from other geographic locations in the United States have found seasonal fluctuations in antibody prevalence over a 10-year period (Calisher et al. 2007). Even with this long-term study, they only detected a clear seasonal pattern in one out of four sites. Part of the problem with detecting seasonal variation is the limitation in trapping during the winter months because of the decrease in Peromyscus population density. Future hantavirus research in Pennsylvania should consist of longer-term studies with increased trapping effort, particularly during early spring and late fall when mouse population density is low. This may provide better temporal resolution and allow better detection of seasonal patterns in prevalence.
In our study, there was a significant male bias in antibody prevalence, with males accounting for approximately three times as many cases as females. This is consistent with several previous studies on hantaviruses (Bennett et al. 1999, Goodin et al. 2009, Luis et al. 2010). Males may be at higher risk of exposure or be more susceptible to hantavirus infection than females. There is some evidence that higher levels of testosterone in males correlate with increased likelihood of hantavirus infection as well as increased shedding of the virus (Klein et al. 2002, Easterbrook et al. 2007). In addition, previous studies have demonstrated that hantavirus antibody-positive mice are more likely to be wounded, suggesting that aggressive encounters may facilitate transmission between Peromyscus (Bennett et al. 1999, Mills et al. 1999, Douglass et al. 2001, Hinson et al. 2004, Calisher et al. 2007, Goodin et al. 2009). Although we did not detect a direct correlation between wounding and antibody prevalence, both wounding and incidence of infection were positively associated with host mass class (proxy for age). Given that antibodies take on average 2 weeks to develop after infection, wounding at the time of antibody detection may not be a reliable gauge for a transmission event as wounds at the time may have healed (Botten et al. 2000). Nevertheless, our results suggest a potentially higher risk of hantavirus transmission in older male animals displaying greater aggression.
In conclusion, our study revealed that one or more hantaviruses are present and circulating in Peromyscus populations in central Pennsylvania. We found spatial and temporal variation in antibody prevalence, which could not be accounted for by variation in host abundance. We investigated host factors likely associated with rodent-to-rodent transmission risk, and found that host sex and age (i.e., mass class) serve as reliable predictors of risk. Older mice were more likely to be wounded, suggesting a relationship between aggression and transmission. This hypothesis is supported by the positive association we detected between wounding and age, and wounding and incidence of infection (sero-conversions). Although our study did not identify the strain of Hantavirus present, the two most likely candidates are New York virus, associated with P. leucopus from several areas of New York State, or Monongahela virus, a subtype of SNV associated with Peromyscus spp. Additional molecular diagnostic work is needed to delineate the species circulating in Pennsylvania. Future ecological studies on hantavirus associations with reservoir hosts will help inform the development of models to predict future outbreaks and protect against zoonotic transmission to humans.
Footnotes
Acknowledgments
We would like to thank K. Vandegrift, D. Grear, and J. Sinclair for collecting the field data and blood samples. We especially thank H. Mays and the Cincinnati Museum Center for assistance with the Peromyscus DNA barcoding. This research was funded by the National Science Foundation (Grant number: 0520468). P.J.H. was supported by the RAPIDD program from NIH.
Disclosure Statement
No competing financial interests exist.