
Abstract
Some foodborne pathogens can cause mastitis, in which the organism is directly excreted into milk. Therefore, we undertook the steps to determine the prevalence and molecular characteristics of Shiga toxin-producing Escherichia coli (STEC) isolates from bovine mastitic milk in Egypt. Forty milk samples from dairy cattle showing mastitis were collected and examined for the presence of E. coli. Following enrichment and plating on selective agar, confirmation of the isolates was based on biochemical tests and the isolates were determined at the species level using cytochrome oxidase, triple sugar iron agar, urea, and indole tests as putatively E. coli. About 77.4% of the isolates belonged to four different O serogroups (O26, O86, O111, and O127). The multiplex polymerase chain reaction (PCR) found that the seven isolates revealed positive amplification of the Eagg gene from the extracted DNA of the E. coli isolates in an incidence of 100%. Also, the selected isolates were subjected to a simple PCR for the detection of 12 of the most important E. coli genes associated with virulence. Those genes detected were stx1, stx2, hylA, Flich7, stb, F41, K99, sta, F17, LT-I, LT-II, and eaeA. A total of seven E. coli isolates that were non-O157 isolates were investigated. Among the seven isolates, none was stx positive, and all seven lacked F41, K99, LT-I, LT-II, and Flich7 . Of these seven isolates, three (42.85%) were enterohemorrhagic E. coli hlyA positive and two (28.57%) were eaeA positive. STEC isolates were not found in bovine mastitic milk in Egypt. Isolates from mastitic milk were potentially pathogenic for human in that they belonged to serogroups associated with diarrhea and hemolytic-uremic syndrome, and some of them were hylA, stb, sta, F17, and eaeA positive.
Introduction
Escherichia coli is considered to be an opportunistic pathogen and originates from a contaminated environment (Nemeth et al. 1994). Consequently, E. coli can cause clonal persistent intramammary infection in dairy cows (Dogan et al. 2006). E. coli is one of the major agents of clinical bovine mastitis worldwide (Shpigel et al. 1998, Longo et al. 2001, Bradley 2002, Burvenich et al. 2003, Hogan and Smith 2003, Blum et al. 2008) and it is an increasing problem. Bovine mastitis caused by E. coli has been traditionally viewed as a transient infection (Dogan et al. 2006). The results support the conclusion that mastitis-causing E. coli bacteria are typical commensals (Suojala et al. 2011). The reason for the importance of E. coli mastitis is its increasing incidence and severe symptoms (Fang and Pyörälä 1996).
Bovine mastitis E. coli do not have specific antigen O serogroups (Saran-Rosenzuaig and Cohen 1972, Sanchez-Carlo et al. 1984b, Lipman et al. 1995a, Correa and Marin 2002, Bean et al. 2004, Wenz et al. 2006, Blum et al. 2008) and are not biochemically different from fecal E. coli (Nemeth et al. 1994). Serotypes O26, O111, O103, and especially O157 have been the predominantly isolated enterohemorrhagic E. coli (EHEC) (Nataro and Kaper 1998). Murinda et al. (2004b) isolated O26, O111, O103, and O157:H7 EHEC serotypes from dairy cows and/or the dairy farm environment. It appeared to Murinda et al. (2004b) that, these serotypes could be of epidemiological significance in the cause of mastitis in the United States. Consequently, we hypothesized that their role in Egypt must not be underestimated.
Bacteria require virulence factors to colonize, multiply, and survive in the udder. Different virulence factors have been already identified in E. coli isolates from bovine mastitis (Nemeth et al. 1991, 1994, Lipman et al. 1995b, Kaipainen et al. 2002, Bean et al. 2004, Lira et al. 2004, Wenz et al. 2006). Yet, the results of Wenz et al. (2006) are consistent with previous reports suggesting that E. coli strains causing acute coliform mastitis in dairy cattle do not possess the same specific virulence factors that contribute to clinical disease (Barrow and Hill 1989, Kaipainen et al. 2002, Bean et al. 2004).
Each year, the E. coli strain causes about 100,000 illnesses, 3000 hospitalizations, and 90 deaths in the United States (Marler 2010). Shiga toxin-producing E. coli (STEC) infection may cause a broad disease spectrum in humans (Levine 1987, Karmali 1989, Griffin and Tauxe 1991). E. coli strain that produced a potent toxin cytotoxic for Vero (green monkey kidney) cells were first reported by Konowalchuk and Speirs (1977). The toxin was appropriately named verotoxin. A number of factors account for the virulence of STEC isolates, and the most prominent among these is the Stx group of toxins (Ito et al. 1990, Gyles 1992, Feng and Monday 2000, Wang et al. 2002, Gyles 2007). Based on serological methods and DNA sequence analysis, these Stx toxins have been divided into two major subclasses, Stx1 and Stx2 (Head et al. 1988).
A wide range of STEC serogroups can cause human disease, although the most commonly isolated serogroup from patients infected with STEC are notably O26, O103, O111, O145, and O157 (Schmidt et al. 1999, Willshaw et al. 2001, Brett et al. 2003). The virulence of the STEC strains is affected by the enterohemolysin (hly) encoded by the hlyA gene (Schmidt et al. 1995) and the eaeA gene, which encodes intimin and is involved in intimate attachment of the bacteria to intestinal host cells (Frankel et al. 1998). However, as STEC non-O157 are more prevalent in animals and as contaminants of food, humans are probably more exposed to these strains, and some of them have been associated with severe illness in humans (Blanco et al. 2003, Lira et al. 2004).
The present study was therefore designed to investigate pathogenic E. coli isolates from bovine mastitic milk for the presence of specific virulence markers and to examine the relationship between serotype and virulence genes of E. coli mastitis isolates that can pose serious health risks.
Materials and Methods
Bacterial strains
Milk samples from cows with mastitis (n=40) were asseptically obtained by veterinarians with laboratory training. Approximately 5 mL of milk was collected in sterile glass bottles, stored on ice, and transported to the laboratory for culture. CM was identified by the California mastitis test (Schalm and Noorlander 1957) and clinical examination. Samples were cultured in MacConkey (MAC) medium. Agar plates were incubated at 37°C and bacterial growth was evaluated after 24 and 48 h, respectively. Gram-negative microorganisms were isolated from MAC agar and determined at the species level using cytochrome oxidase, triple sugar iron agar, urea, and indole tests as putatively E. coli (Quinn and Markey 2003). Reference strains used were E. coli strain ATCC 35150 (O157:H7, stx1, stx2, eae, hly) (positive control) and Staphylococcus aureus ATCC 29737 (negative control) (Chart et al. 2000).
Detection of O-serogroups
E. coli isolated in MAC were biochemically confirmed and submitted to slide agglutination tests using polyvalent and monovalent sera against serogroups O26, O25, O86, O111, O78, O119, O126, O127, O164, O157, and O158. Commercially available antisera in the Central Laboratories of Ministry of Public Health, Egypt, were used.
Hemolytic activity
The strains were tested for h-hemolytic activity on agar base (Oxoid) supplemented with 5% sheep erythrocytes. Five microliters of each suspension was streaked onto the plates, and the plates were incubated at 22°C and 37°C for 24 h. The presence of a clear colorless zone surrounding the colonies indicated h-hemolytic activity (Gerhard et al. 1981).
Congo red dye uptake
The ability to take up Congo red dye was determined on agar plates supplemented with 50 Ag/mL of Congo red dye. Five microliters of each suspension was streaked onto the plates, and the plates were incubated at 37°C for 24 h. Orange colonies were considered positive; different intensities in the dye uptake were expressed as +, ++, and +++ (Paniagua et al. 1990).
Vero cell cytotoxicity
E. coli isolates were grown in brain heart infusion broth for 8 h at 41°C. Then, 5 mL of each isolate was subcultured into 50 mL Casamino acid–yeast extract–salts medium without glucose. The cells were allowed to aerobically grow at 37°C. After 18–20 h, the cells were removed by centrifugation at 12,000 g for 15 min at 4°C. The cell pellet was resuspended in 0.5 mL sterile phosphate-buffered saline (0.1 M, pH 7.2). The cells were ultrasonically disrupted continuously for 2 min in an ice bath using a sonicator. The cytotoxicity assay was performed with Vero cells as previously described (Emery et al. 1992).
DNA isolation
E. coli strains were subcultured from frozen or peptone stocks onto MAC agar plates, utilizing quadrant streaking methods to produce isolated colonies. These strains were then placed in an incubator for culture at 37°C. After overnight incubation, a single colony was carefully removed from the plate by using a sterile toothpick to avoid agar contamination, an important cause of erratic amplification. Crude lysates were prepared and directly used as a template for the polymerase chain reaction (PCR) (Sritharan and Barker 1991). DNA was extracted by boiling a single colony in 50 μL of PCR- or molecular-grade water for 5 min, followed by centrifugation at 14,000 rpm for 10 min. Two microliters of this lysate was used as a template with a 23 μL PCR master mix to make a 25 μL total reaction volume.
DNA amplification of the Eagg and Einv genes of E. coli
The amplified reactions were performed in 50 μL volumes in microamplification tubes (PCR tubes). The primers and multiplex-PCR conditions are described in Table 1. The reaction mixture consisted of 10 μL (200 ng) of extracted DNA template from bacterial cultures, 5 μL of 10× PCR buffer, 0.75 μL MgCl2 (3 mM), 0.5 μL dNTPs (100 μM), 0.5 μL (2.5 U) Ampli Taq DNA polymerase, and 0.1 μL (0.2 Pmol/μL) from each primer pair, and the volume of the reaction mixture was completed to 50 μL using double-distilled water (DDW).
Unless stated, PCR was done for 25 cycles.
After 20 cycles, final extension step of 5 min at 72°C was performed.
After 35 cycles, final extension step of 10 min at 72°C was performed.
After 25 cycles, final extension step of 10 min at 70°C was performed.
PCR, polymerase chain reaction.
Detection of stx1, stx2, hylA, Flich7, stb, F41, K99, sta, F17, LT-I, LT-II, and eae sequences
The E. coli isolates were subjected to simplex-PCR, performed in an Eppendorf Mastercycler (Ependorf AG). The stx1, stx2, hylA, Flich7, stb, F41, K99, sta, F17, LT-I, LT-II, and eaeA genes were detected using the primers and s-PCR conditions described in Table 1. An initial denaturation step at 95°C for 3 min (stx1, stx2, hylA, Flich7, stb, F41, K99, sta, LT-I, LT-II, and eaeA genes) and 94°C for 5 min (F17) was carried out in separate steps for each gene. The s-PCR mixtures fell into five sets:
Set A
An amount of 0.1 μL (0.2 μM concentration) of each of the primers for stx1, stx2, hylA, Flich7 , and stb, in separate analyses, together with 1.25 μL (250 μM) deoxynucleoside triphosphates, 5.0 μL of 10× reaction buffer, 0.1 μL (0.5 U) of Taq DNA polymerase, and 10 μL (200 ng) of template DNA was taken. The volume of each mix was adjusted to 50 μL with DDW.
Set B
An amount of 0.25 μL (0.5 μM) of each of the primers for the detection of F41 and K99 genes together with 1.25 μL (250 μM) deoxynucleoside triphosphates, 5.0 μL of 10× reaction buffer, 0.25 μL (1.25 U) of Taq DNA polymerase, and 10 μL (200 ng) of template DNA was prepared. The volume of each mix was adjusted to 50 μL with DDW.
Set C
An amount of 1.25 μL (250 μM) deoxynucleoside triphosphates, 5.0 μL of 10× reaction buffer plus each primer added separately for detection of the F17 fimbriae (P7, P8, and P9 adhesin genes) at a concentration of 0.6 μM of each of the primers together with 0.25 μL (1.25 U) of Taq DNA polymerase and 10 μL (200 ng) of template DNA was prepared. The volume of each mix was adjusted to 50 μL with DDW.
Set D
An amount of 1.25 μL (250 μM) deoxynucleoside triphosphates, 5.0 μL of 10× reaction buffer in addition to 0.1 μL of each of the primers separately of the LT-I and LT-II genes at a concentration of 0.2 pmol of each of the primers together with 0.5 μL (2.5 U) of Taq DNA polymerase and 10 μL (200 ng) of template DNA was prepared. The volume of each mix was adjusted to 50 μL with DDW.
Set E
An amount of 0.5 μL (100 μM) deoxynucleoside triphosphates, 5.0 μL of 10× PCR buffer in addition to the eae gene primer at a concentration of 0.25 μL (0.5 μM) together with 0.25 μL (1.25 U) of Taq DNA polymerase and 10 μL (200 ng) of template DNA was prepared. The volume of each mix was adjusted to 50 μL with DDW.
The PCR products were observed after electrophoresis on 1.2% agarose gel and staining with ethidium bromide. The results were recorded using an Alpha Imager documentation camera.
Results
Among the eight E. coli isolates analyzed, the somatic antigen (serogroup) was determined in seven isolates. A total of 77.4% of the isolates belonged to four different O serogroups (O26, O86, O111, and O127) and one isolate belonged to a nonidentified serogroup (Table 2).
UD, undetected.
The seven isolates were not able to produce cytopathic effect on the Vero cells, although they were 100% hemolytic.
Congo red assay was used as a phenotypic marker for the invasive and noninvasive E. coli. In the present study, all of the seven tested serovars for the Congo red (CR) binding affinities were 100% positive. The binding activity of the CR dye was found to be variable in their affinity according to their serovars.
The results of the m-PCR as observed in Table 3 and Figure 1 revealed a negative result for the amplification of the 140-bp fragment for the Einv gene in the seven field isolates in an incidence of 100%. On the other hand, the 28 field isolates revealed positive amplification for the 194-bp fragment of the Eagg gene from the extracted DNA of the E. coli isolates in an incidence of 100%.
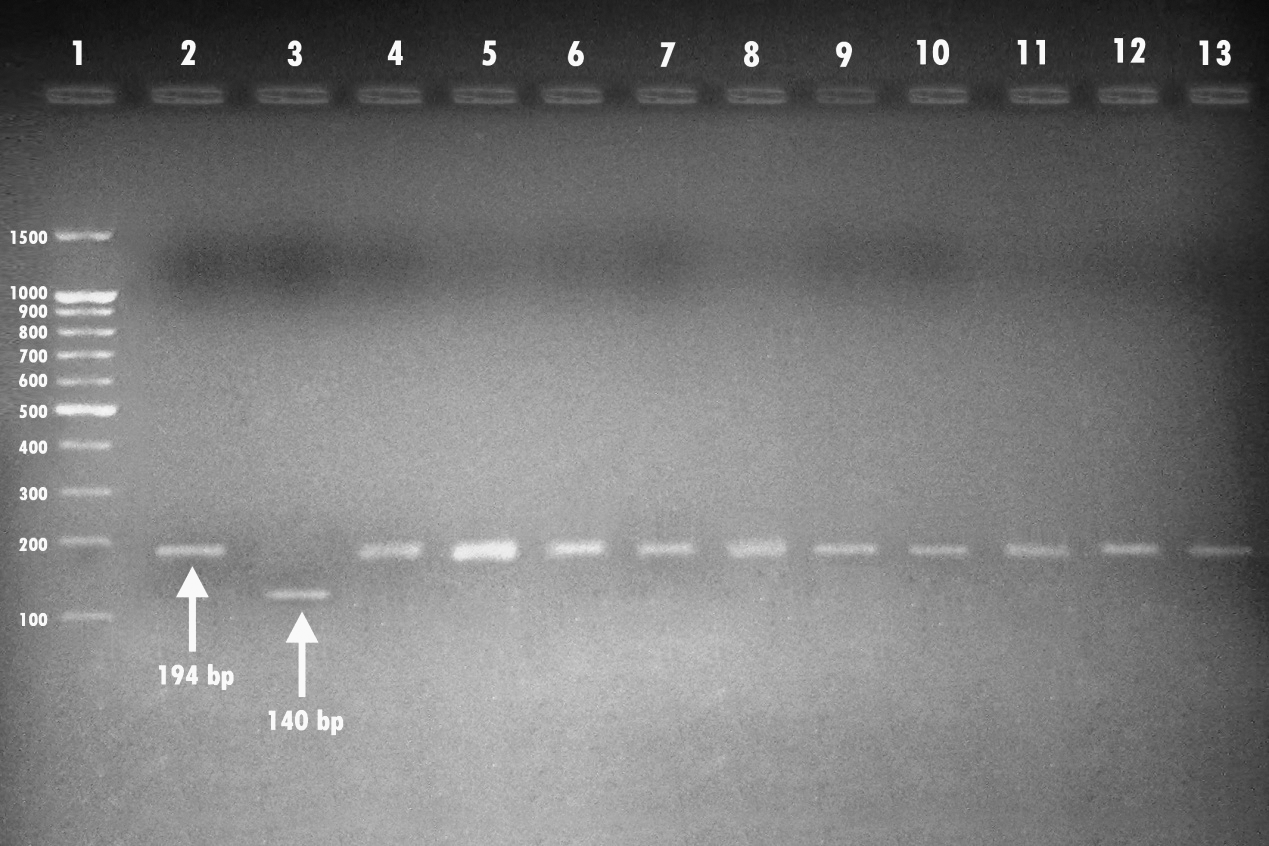
Agarose gel electrophoresis showing amplification of the 194-bp and 140-bp fragments for the Eagg and Einv genes of Escherichia coli using the multiplex-polymerase chain reaction performed with their specific primers. Lane 1, 100-bp-molecular-weight ladder; lane 2, control E. coli HB 101 inv+. lane 3, positive control (E. coli ATCC 35150 serotype O157:H7); lanes 4–13 show positive amplification of the 194-bp fragment. All of the bacterial strains showed positive amplification for the Eagg gene of E. coli.
The distribution pattern of the detected virulence genes was observed to be in the following order: F17 (57.14%), Sta (42.85%), hylA (42.85%), eaeA (28.57%), and Stb (14.28%). The rest of the assayed genes not detected were stx1, stx2, F41, K99, LT-I, LT-II, and Flich7 (Table 4).
Seven (100.0%) of the isolates carried the F17 gene, predominantly associated with eaeA and Sta genes. Three (42.85%) of the isolates were positive for Stb and F17 genes in conjunction. Two (28.57%) of the isolates were detected carrying both of the eaeA and hylA genes in combination. Only one isolate carried the hylA gene alone (14.28%) (Table 5).
The seven (17.5%) isolates contained genes encoding for serotypes O26:K60, O86:K61, O111:K58, and O127:K-LPS, which are more frequently associated with human infections. The gene eaeA encoded serogroups O26:K60 (one isolate; 14.28%) and O127:K- (one isolate; 14.28%); hylA encoded serogroups O86:K61 (one isolate; 14.28%), O111:K58 (one isolate; 14.28%), and O127:K- (one isolate; 14.28%); Sta encoded serogroup O26:K60 (three isolates; 42.85%); Stb encoded serogroup O111:K58 (one isolate; 14.28%); and F17 encoded serogroups O26:K60 (three isolates; 42.85%) and O111:K58 (one isolate; 14.28%) (Table 6).
Discussion
E. coli, a gram-negative environmental pathogen associated with bovine mastitis, was isolated from the milk of 40 symptomatic cows that had been diagnosed with CM. In France, E. coli (16%) was identified as one of the major causative agents of CM (Botrel et al. 2010), a prevalence that is very close to our findings in Egypt (17.5%).
Several authors have reported a wide range of serogroups in cattle (Wells et al. 1991, Wieler et al. 1996, Holland et al. 1999, Correa and Marin 2002, Wani et al. 2003); some of them were found only in bovine and others such as O26, O103, O111, and O157 have been found in bovine and humans (Paton and Paton 1998, Bettelheim 2000). More than 100 different STEC serotypes have been isolated from cattle, although some serotypes are more frequently isolated than others (Beutin et al. 1993). In the present investigation, the serogroup O26:K60 (three isolates) was more prevalent than serogroups O86:K61, O127:K-, and O111:K58, which are frequently associated with human infections. In a study conducted by Wenz et al. (2006), no predominant serotype was identified by farm or systemic disease severity; however, the most frequent serotype was O158:NM (n=3). In cattle, there is often a continual turnover of serotypes of E. coli (Hinton et al. 1985).
Tissue culture cytotoxicity assays play an important role in detecting E. coli O157 and other STEC organisms in samples from which isolation of these organisms is difficult. Consequently, it was appropriate to use it in our investigation. Konowalchuk and Speirs (1977) first observed that Vero cells possess profound sensitivity to Shiga toxin (STX), and up to the present time, this cell line remains the “gold standard” for confirmation of the cytotoxicity of putative S-producing organisms. Vero cells possess a high concentration of both Gb3 and Gb4 receptors on the surface of their plasma membranes, which makes this cell line ideal to detect all known STX variants. The assay involves treatment of Vero cells at the confluent stage with sterile culture extracts and observing cells for a cytopathic effect after 48–72 h of incubation (Karmali 1989). The results were consistent with the PCR assay in that the STX was not detected in our isolates.
Reports on the prevalence of STEC associated with mastitis are rare. No Shiga toxin in E. coli isolates from mastitis was reported in some studies (Lipman et al. 1995a, Cullor, 1997, Murinda et al. 2004a, Guler and Gunduz 2007), suggesting that alternative virulence characteristics are involved in mastitis. Nevertheless, in some other studies, STEC genes have been found from bovine mastitis (Barrow and Hill 1989, Stephan and Kuhn 1999, Lira et al. 2004, Vu-Khac and Cornick 2008). The most common virulence gene detected was stx1, with a prevalence of 31%, followed by eaeA (4%) (Bean et al. 2004, Murinda et al. 2004a, Wenz et al. 2006).
Thus, it would appear that STEC are rarely associated as a causative agent of bovine mastitis. In the study conducted by Guler and Gunduz (2007), 9% of the strains were positive for the F17 fimbriae, a percentage significantly lower than that in our results (57.14%). The intimin gene was found in one strain by Guler and Gunduz (2007), in three strains by Vu-Khac and Cornick (2008), and in two isolates by us.
Enterotoxin production is also not a common property of E. coli isolates from mastitis. Barrow and Hill (1989) detected enterotoxin in only 1 of 237 isolates. Sanchez-Carlo et al. (1984a) found only 1 heat-stable enterotoxin and 2 heat-labile enterotoxin positive isolates among 184 E. coli isolates from cows with acute mastitis. Lipman et al. (1995a) found no strains that were positive for enterotoxin genes (LT and ST1) among 20 isolates. Similarly, in the study by Guler and Gunduz (2007), no heat-stable enterotoxin gene was detected. Our results indicated that the isolates were negative to LT-I and LT-II heat-labile enterotoxin, which agrees with Lipman et al. (1995a), and positive for the production of heat-stable enterotoxin (Sta and Stb), which does not agree with Guler and Gunduz (2007). Dogan et al. (2006) found no evidence of the K99, LT, Sta, Stb (associated with ETEC), stx1 and stx2 (associated with EHEC), hly, Sta (associated with UPEC), and eaeA (associated with EPEC and EHEC) virulence genes typically associated with invasion of diarrheagenic and uropathogenic E. coli to account for the invasive behavior of mastitis-associated E. coli strains. This result contrasts with reports showing that the majority of mastitis-associated E. coli is consistent with the absence of the virulence gene eaeA (Bean et al. 2004, Dogan et al. 2006, Vu-Khac and Cornick 2008).
Kaipainen et al. (2002) found that 49% of Finnish and 42% of Israeli isolates had at least one virulence gene, which is significantly higher than our results (17.5%). We also found that F17 was the most common gene found in 57.14% of our isolates, whereas it was 9% in the Finnish isolates, 1% of the Israeli isolates (Kaipainen et al. 2002), and 20.47% in Iran (Ghanbarpour and Oswald 2010). Among the seven E. coli isolates, 28.57% had eaeA, 42.85% Sta, and 14.28% Stb genes. Also, we detected the hlyA gene (42.85%), but Suojala et al. (2011) failed to detect the gene. In the present study, it was observed that all of the detected virulence genes were present alone or in combination as previously observed by Ghanbarpour and Oswald (2010). The absence of virulence genes Stx1, Stx2, Flich7, F41, K99, LT-I, and LT-II coincides with findings of previous authors (Bean et al. 2004, Murinda et al. 2004a, Dogan et al. 2006, Suojala et al. 2011).
The STEC strains seem to be pathogenic for humans only if they possess accessory virulence factors (Lira et al. 2004). The eae gene is responsible for attachment and effacement lesions similar to those in enteropathogenic E. coli (Gannon et al. 1993). In most human STEC strains belonging to enterohemorrhagic serotypes, eae genes are present. In this study, two (28.57%%) of the isolates were positive for eaeA gene, which is more frequently found in cattle with diarrhea (Blanco et al. 1997) or healthy cattle (Osek 2002). Despite the claim that the eae gene is a major marker of virulence, there is evidence that STEC strains causing severe human disease do not necessarily contain the eaeA gene (Dytoc et al. 1994). A comparison of eae-positive and eae-negative STEC isolates in this study showed a linkage between eaeA and hly (28.57%), which agrees with the results of Sandhu et al. (1996).
The discrepancies found in the differences in the proportion of mastitis caused by E. coli, clinical pictures of coliform mastitis, serogroups, the distribution pattern of the detected virulence genes, and selection and combination of virulence genes could be attributed to environmental conditions, geographical location, season, farm size, number of animals on the farm, hygiene, farm management practices, variation in sampling, variation in types of samples evaluated, and differences in detection methodologies used (Kaipainen et al. 2002, Oliver et al. 2005).
The results indicated that there are no antigen serogroups host specific. Also, these studies indicate that there was no specific virulence factor that is commonly produced by E. coli isolates from ovine and bovine mastitis and diarrhea.
Taking into consideration that E. coli from mastitis originates from fecal flora or the cow's environment, virulence characteristics of the isolates from mastitis in this study may also reflect the general situation of bacteria in the environment and their propagation through environmental vectors. Flies transmit diseases including bacterial mastitis infections (Anonymous 2010). Mastitis bacteria can live up to 16 h in a fly, which is more than enough time to transmit the bacteria from cow to cow. Studies by Sanders (1940) have incriminated the Musca domestica (housefly), the genus Hippelates (fruit fly or eye gnat) (Anonymous 1940), and the headfly Hydrotaea irritans (Fallén) (Madsen et al. 1992) as vectors of bovine mastitis.
Therefore, supporters of tail docking claim that tail docking reduces udder hygiene, increasing the chances for mastitis (Admin 2009). The transmission of disease among cows by external parasites is yet another potential cause for reduced milk production and mastitis.
Footnotes
Disclosure Statement
No competing financial interests exist.