
Abstract
Fleas were collected from live-captured small mammals to identify flea-borne pathogens, host associations, and seasonal prevalence of flea species, as part of the 65th Medical Brigade rodent-borne disease surveillance program at 20 military installations and training sites, Gyeonggi Province, Republic of Korea, 2005–2007. A total of 1251 fleas were recovered from 2833 small mammals. Apodemus agrarius, the striped field mouse, accounted for 93.1% (2,637/2,833) of all small mammals captured, followed by Crocidura lasiura (3.1%), Mus musculus (1.3%), Microtus fortis (0.7%), Myodes regulus (0.7%), Micromys minutus (0.5%), Rattus norvegicus (0.4%), Tscherskia triton (0.1%), Apodemus peninsulae (<0.1%), Rattus rattus (<0.1%), and Mogera robusta (<0.1%). A total of 6/11 species of mammals captured were infested with fleas with infestation rates ranging from a high of 26.3% (A. agrarius and M. regulus) to a low of 5.3% (M. fortis). Flea indices among infested mammals were highest for R. norvegicus (2.50), followed by C. lasiura (2.20), A. agrarius (1.71), M. regulus (1.20), M. musculus (1.0), and M. fortis (1.0). The predominant flea species collected were Stenoponia sidimi (56.5%), followed by Ctenophthalmus congeneroides (38.3%) and Rhadinopsylla insolita (3.9%). The minimum field infection rates [number of positive pools/total number of fleas (600)] for Rickettsia typhi and for Rickettsia felis were 1.7% and 1.0%, respectively.
Introduction
The first recorded survey of rat fleas in Korea was conducted by Kobayashi in 1931, with subsequent surveys reporting a total of 37 flea species and associated host relationships (Nagahana 1954, Tipton et al. 1972, Hong 1994). After the systematic identification of ectoparasites and host relationships, zoonotic pathogens were identified in rodents and associated hosts (Kim et al. 2003, 2005, 2006, Chae et al. 2008, Kim et al. 2010b, O'Guinn et al. 2010, Sames et al. 2010). The development of rodent-borne disease surveillance programs, which identify the potential for human exposure to vector-borne disease agents observed in feral and domestic animals as well as ectoparasite diversity, provide a better understanding of disease maintenance cycles. In turn, this information can be used to identify and predict disease emergence events (Nieto et al. 2007).
Flea species, and associated hosts and pathogens were identified to determine the prevalence of the Rickettsia felis and R. typhi in fleas collected from small mammals in the ROK. In addition, the relative seasonal abundance of fleas collected from wild-caught small mammals at military training sites was determined. These data are useful for compiling vector-borne disease risk assessments for pathogens transmitted by fleas at military training sites and installations in the ROK.
Materials and Methods
Samples
Fleas were collected from small mammals that were live-trapped at 4 U.S. military installations, 15 U.S. and ROK–operated military training sites, and 1 communications post, in Gyeonggi Province, ROK (Fig. 1). Surveys were conducted seasonally from 2005 to 2007 during the spring (March–April), summer (June), fall (late August–September), and winter (late November–December). Sherman traps (7.7×9×23 cm aluminum collapsible live-traps (H.B. Sherman, Tallahassee, FL), baited with peanut butter placed between two saltine crackers, were set out during daylight hours and collected the following morning. Since nighttime temperatures often fell below 0°C, nonabsorbent cotton balls were placed in each trap during the winter and spring trapping periods so that the trapped animals would retain heat until processed. Live small mammals were transported to Korea University and euthanized under an approved animal use protocol as described by O'Guinn et al. (2010). Fleas were removed, identified to species using conventional taxonomic keys, then placed in 1.0 mL cryovials, and stored individually at −70°C until assayed for selected pathogens (Hopkins and Rothschild 1953, 1956, Hong 1994). Flea infestation rates, indices, and minimum field infection rates (MFIR) were determined by the following formulas:
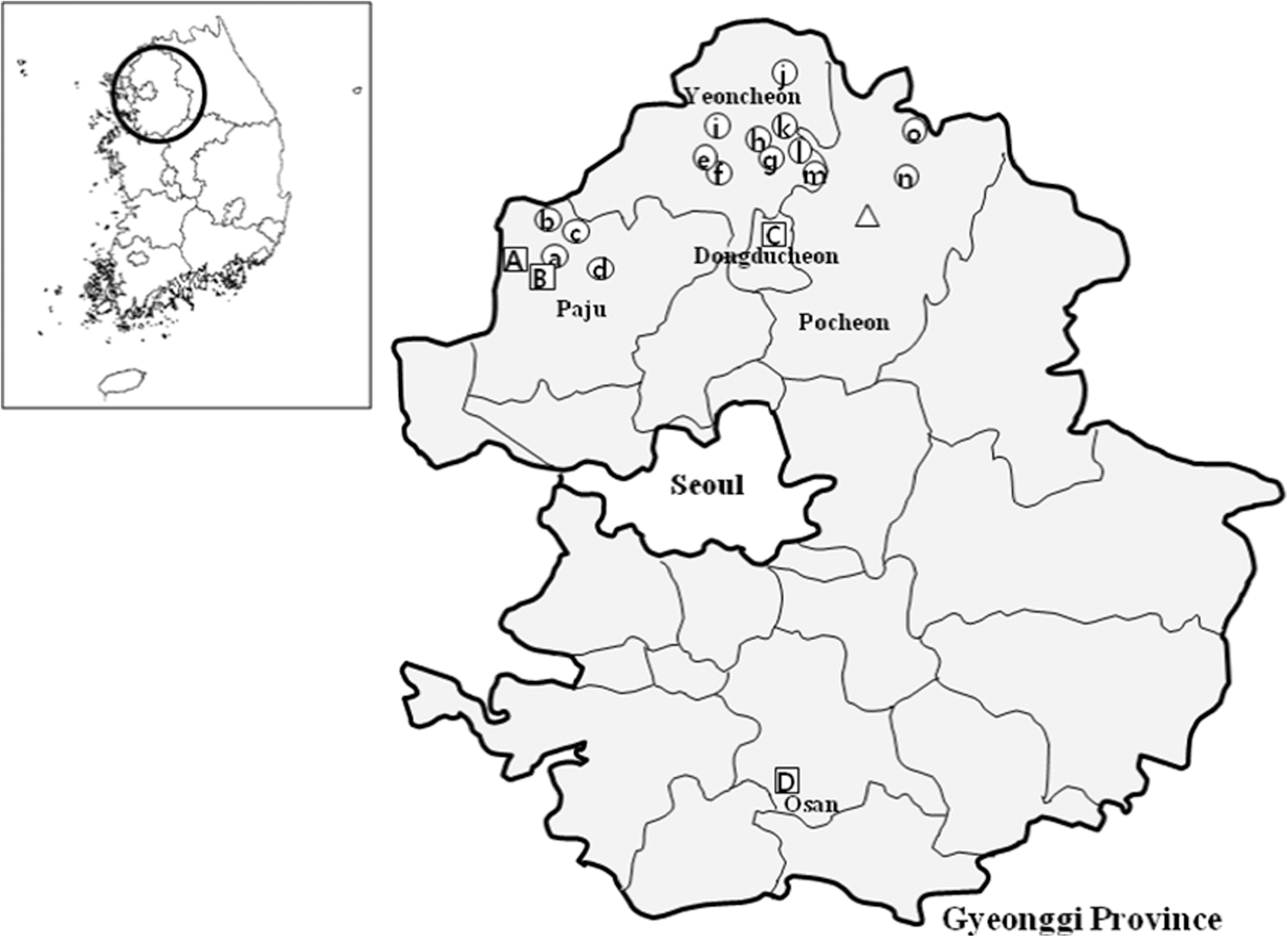
Geographical locations of small mammal collection sites at 15 U.S. and ROK–operated training sites (○), 4 U.S. military installations (□), and 1 Communications Post (▵), Gyeonggi Province, ROK: Camp Bonifas (A) and Warrior Base (B), Gunnae-myeon, Paju-si; Camp Casey (C), Bosan-dong, Dongducheon-si; Osan AB (D), Osan-si; KC-39 (Δ), Pocheon-gun; Monkey Range 7(a) and Story Range(b), Jindong-myeon, Paju-si; Dagmar North Training Area (c), Jeokseong-myeon, Paju-si; Twin Bridge Training Area (d), Jikcheon-ri, Beobwon-eup, Paju-si; Local Training Area [LTA 130 (e), 320 (f), 36(g), 37(h)], and Chaparral Training Area (i), Misan-myeon, Yoencheon-gun; Santa Barbara Range (j), Yeoncheon-gun; Firing Point (FP) 10(k), 60 (l) and 66 (m), Yeoncheon-gun; Rodriguez Range (n), Youngjung-myeon, Pocheon-gun; and Nightmare Range (o), E-dong, Pocheon-si, Gyeonggi Province, ROK.
Flea Infestation Rate=Number of Captured Small Mammals with Fleas/Total Number of Small Mammals Captured
Flea Index=Number of Fleas Collected from Small Mammals/Number of Small Mammals Infested
MFIR*=Number of Positive Pools of Fleas/Total Number of Fleas
*Based on a maximum of one positive (infected) flea per positive pool, although there may be more than one positive flea in the pool.
DNA extraction
DNA was extracted from 300 pools (600 fleas, 2 fleas/pool) for detection of R. felis and R. typhi. Fleas were pooled according to collection dates and sites, flea species, and host animal species. Fleas were homogenized mechanically using a Beadbeater TissueLyser II (QIAGEN, Hilden, Germany) with 180 μL lysis buffer, 20 μL proteinase K (600 AU/mL), and 5 mm stainless steel beads at 30 frequencies/s for 5 min, followed by incubation at 56°C overnight and centrifugation at 12,000 g for 15 min at room temperature. After centrifugation, the supernatant was used for genomic DNA extraction performed with DNeasy® Tissue Kits (QIAGEN) according to the manufacturer's instructions.
Polymerase chain reaction detection
Extracted DNA was amplified in two separate nested-polymerase chain reaction (PCR) runs with different target genes to confirm results and differentiate Rickettsia species. All samples were first screened using genus-specific primers (RpCS.877p: 5′-GGGGACCTGCTCACGGCGG; RpCS.1258n: ATTGCAAAAAGTACAGTGAACA; RpCS.896p: GGCTAATGAAGCAGTGATAA; RpCS.1233n: GCGACGGTATACCCATAGC), derived from the citrate synthase–encoding (gltA) gene for Rickettsia (Roux et al. 1997). Screened samples that were positive were selected for a second PCR assay with species-specific primers targeting the ompB gene (rompB OF: GTAACCGGAAGTAATCGTTTCGTAA, rompB OR: GCTTTAATACGTGCTGCTAACCAA, rompB SFG IF: GTTTAATACGTGCTGCTAACCAA, rompB SFG/TG IR: GGTTTGGCCCATATACCATAAG) (Choi et al. 2005).
Positive controls for R. typhi (strain Wilmington) and R. felis (strain URRWXCal2) DNA were provided by the Division of Zoonosis, Center for Immunology and Pathology, Korea National Institute of Health, and the Viral and Rickettsial Diseases Department, Naval Medical Research Center, respectively. To minimize the possibility of nested product contamination, two separated rooms, which had dedicated PCR equipment, supplies, and solutions, were used. Negative controls, including a water control, negative wild rodent tissues, and flea DNA, were included for each of the nested PCR assays. No detectable bands representing R. typhi or R. felis were observed from negative controls.
DNA sequencing and phylogenetic analysis
The PCR products were purified with QIAquick Gel Extraction kits (QIAGEN). After purification, all 16 gltA-positive PCR products were cloned with pGEM®-T Easy Vectors (Promega Corporation, Madison, WI), followed by transformation into Escherichia coli JM109, and then plated onto LB agar containing 50 μg/mL of ampicillin. Three clones for each of the amplicon plasmid DNA for sequencing were purified using the Wizard® Plus SV Minipreps DNA Purification System (Promega Corporation) according to the manufacturer's instructions. Purified recombinant plasmid DNA was sequenced using a T7 and SP6 promoter primer set by dideoxy termination with an automatic sequencer (ABI 3730xl capillary DNA sequencer, NY). Comparative analyses of the nucleotide sequences were completed using Rickettsia gltA gene sequences in the GenBank database. Otherwise, all 12 positive amplicons of ompB gene fragments were sequenced directly. Phylogenetic analyses of the gltA and ompB gene fragments were constructed using the ClustalX 1.60 program and a phylogenetic tree deduced by the neighbor-joining method with MEGA 4.0 software. Numbers on the branches represent bootstrap support (>50%) generated from 500 replications.
Results
From March 2005 through December 2007, a total of 2833 small mammals belonging to seven genera of rodents, one shrew genus, and one mole genus were collected (Table 1, Fig. 1). Apodemus agrarius, the striped field mouse, accounted for 93.1% (2,637) of the small mammals captured, followed by Crocidura lasiura (3.1%), Mus musculus (1.3%), Microtus fortis (0.7%), Myodes regulus (0.7%), Micromys minutus (0.5%), Rattus norvegicus (0.4%), and Tscherskia triton (0.1%). An additional three species (accounted for <1.0% of the total number of small mammals captured (Table 1).
Percent of small mammals captured with fleas.
Average number of fleas per infested animal for each species.
Flea index (FI)=Numbers of fleas/total numbers of infested rodents.
A total of 1251 fleas (869 females and 382 males) were collected from 725/2,833 (25.6%) small mammals. Small mammal infestation rates for 6/11 species of small mammals infested with fleas ranged from a low of 5.3% (M. fortis) to a high of 26.3% (A. agrarius and M. regulus). The overall flea index for all flea infested mammals was 1.72. Flea indices were highest for R. norvegicus (2.50), followed by C. lasiura, (2.20), A. agrarius (1.71), M. regulus (1.20), M. musculus (1.0), and M. fortis (1.0) (Table 1).
Seasonal surveys were conducted during 2005, whereas surveys were only conducted during the winter and spring seasons during 2006 and 2007. For sites that were surveyed during all four seasons during 2005, the overall flea infestation rates were lowest during the spring (13.1%) and highest during the summer season (40.6%). The highest numbers of rodents infested with fleas were found at Rodriguez Range (36.8%), and Dagmar North Training Area (28.8%) (Table 2). For all sites surveyed, the most common flea species recovered was Stenoponia sidimi (56.5%), followed by Ctenophthalmus congeneroides (38.3%), Rhadinopsylla insolita (3.9%), and Neopsylla bidentatiformis (0.4%; Table 3). An additional five species accounted for <1.0% of all fleas collected. Doratopsylla coreana was infrequently encountered and only collected from the Ussuri white-toothed shrew, C. lasiura. While C. congeneroides was collected during all seasons surveyed, the highest populations were observed during the spring through fall seasons. Conversely, S. sidimi was most commonly collected during the winter survey period (Fig. 2). Species R. insolita and S. sidimi were collected only during the spring and winter survey periods, whereas Hystrichopsylla microti and D. coreana were collected only during the winter survey period. Local Training Area 320 and Camp Casey, which were not surveyed seasonally, demonstrated high infestation rates during the December collection period.

Mean number of the two most frequently collected species of fleas of infested small mammals and percent of infested for all flea species captured during all four seasons at nine U.S. and ROK–operated military training sites in northern Gyeonggi Province, ROK, in 2005.
Dagmar North Training Area: Jeokseong-myeon, Paju-si, Gyeonggi province; Camp Bonifas, and Warrior Base: Gunnae-myeon, Paju-si, Gyeonggi province; Monkey Range 7, and Story Range: Jindong-myeon, Paju-si, Gyeonggi province; Firing Point (FP) 10, 60 and 66: Yeoncheon-gun, Gyeonggi province; Chaparral Training Area, and Local Training Area (LTA) 36,37,130, 320: Misan-myeon, Yoencheon-gun, Gyeonggi province; Rodriguez Range: Youngjung-myeon, Pocheon-gun, Gyeonggi province; Twin Bridge Training Area: Jikcheon-ri, Beobwon-eup, Paju-si, Gyeonggi province; Osan AB: Osan-si, Gyeonggi province; KC 39: Pocheon-gun, Gyeonggi province; Camp Casey: Bosan-dong, Dongducheon-si, Gyeonggi province; Nightmare Range: E-dong, Pocheon-si, Gyeonggi province; Santa Barbara Range: Yeoncheon-gun, Gyeonggi province, ROK.
Flea index=Number of fleas/total number of infested rodents.
Number of small mammals captured.
This sample was collected during the winter seasons (Dec) of 2005 and 2006.
ns=not surveyed.
No fleas were collected from Apodemus peninsulae, Micromys minutes, Rattus rattus, Tscherskia triton, or Mogera robusta.
A total of 10/300 pools (600 fleas) were positive for R. typhi (3.3%) with an MFIR of 1.7%, whereas six of the pools were positive for R. felis (2.0%) with an MFIR of 1.0% (Table 4). Sixteen and 12 samples were positive using gltA and ompB gene detecting primers, respectively. Eight gltA nucleotide sequences (2 R. typhi and 6 R. felis) were obtained, whereas sequencing for eight amplicons failed. In the second PCR, 4/16 gltA-positive DNAs were not amplified with ompB gene targeting primers (Table 5). R. felis–positive pools were observed for C. congeneroides, R. insolita, and S. sidimi collected from A. agrarius; from LTA 130 (Yeoncheon), Warrior base (Paju-si), Twin bridge training area (Paju-si), and Camp Casey (Dongducheon-si), Gyeonggi Province (Table 5). A total of 8/10 R. typhi–positive pools were from fleas collected from A. agrarius, whereas the other two positive pools were identified from C. lasiura. The MFIRs for R. typhi were 1.5% for S. sidimi (5/340), 3.2% for R. insolita (2/62), 0.5% for C. congeneroides (1/190), and 50.0% for D. coreana (2/4) (Tables 4 and 5). Acquired ompB oligonucleotide sequences of the PCR products demonstrated 98.7% and 99.3% identity to reference sequences of R. typhi, strain Wilmington (L04661), and R. felis, strain California 2 (AF210695), respectively.
2 fleas per pool (total 600 fleas).
MFIR (Minimum field infection rates)=Number positive pools/Total number fleas assayed.
−, not amplified; +, amplified and not sequenced; ++, amplified and sequenced.
Sequences reported in this article have been deposited in the GenBank database. The phylogenetic tree based on ompB gene sequences [accession numbers: R. felis strain KRS 4 (HQ236989); R. typhi strain KRS 13 (HQ236990)] (Fig. 3) and gltA gene sequences [R. felis strain KRS 1 (JF 448468), KRS 7 (JF 448469), KRS 12 (JF 448470), KRS 11 (JF 448471), KRS 4 (JF 448472), KRS 14 (JF 448473); R. typhi strain KRS 3 (JF 448474), KRS 5 (JF 448475)] (Fig. 4) illustrates the positions of strains identified in this study.
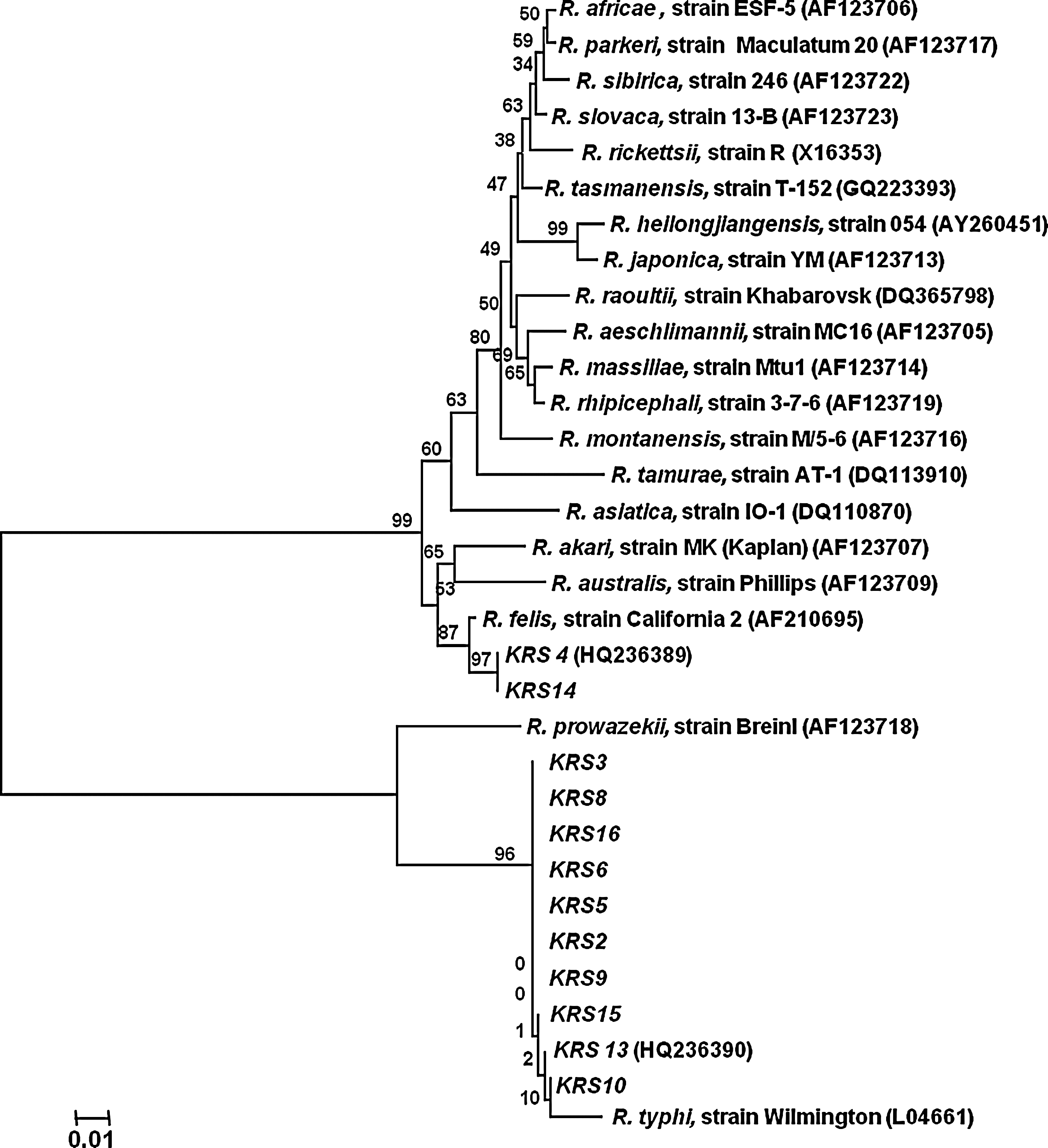
Phylogenetic tree for members of the genus Rickettsia inferred from comparison of ompB sequences using the neighbor-joining method. The phylogenetic tree shows the position of Rickettsia felis (KRS 4, HQ236989) and Rickettsia typhi strains (KRS 13, HQ236990) identified in Gyeonggi Province, ROK, 2005–2007. Bootstrap values for the nodes are indicated.
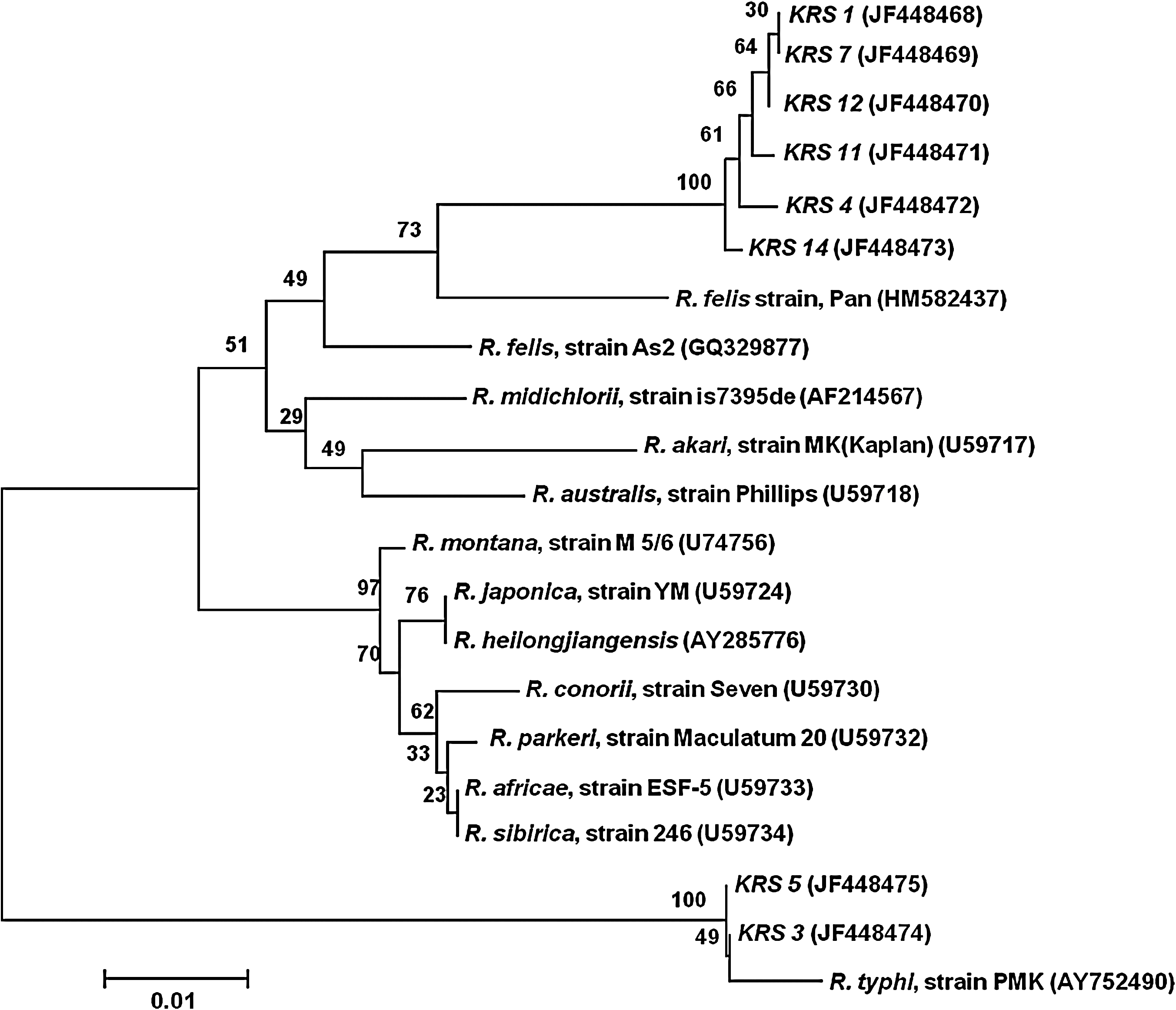
Phylogenetic tree for members of the genus Rickettsia inferred from comparison of gltA sequences using the neighbor-joining method. The phylogenetic tree shows the position of R. felis [KRS 1 (JF 448468), KRS 7 (JF 448469), KRS 12 (JF 448470), KRS 11 (JF 448471), KRS 4 (JF 448472), and KRS 14 (JF 448473)] and R. typhi [KRS 5 (JF 448475) and KRS 3 (JF 448474)] strains identified in Gyeonggi Province, ROK, 2005–2007. Bootstrap values for the nodes are indicated.
Discussion
Adult fleas are mobile obligate parasites that will often abandon their natural host(s), especially after death of the host, and infest alternate hosts, including humans, to obtain blood meals (Azad et al. 1977, Schriefer et al. 1994, Higgins et al. 1996). During 2005, when training sites were surveyed during all seasons, flea infestations were generally higher during the mid-winter months, similar to that observed by Walton and Hong (1976). The exception was C. congeneroides, which was collected more frequently during the spring, summer, and fall survey periods. The biology of these species is not well known in Korea and may reflect host and/or ectoparasite reproductive seasonal differences.
R. felis was identified as a flea-borne rickettsial pathogen in 1990, with C. congeneroides as the primary zoonotic vector playing an important role in human transmission (Adams et al. 1990). Recently, noticeable increases have been observed worldwide in the number of human R. felis infections reported based on PCR or serological tests (Nogueras et al. 2006, Oteo et al. 2006). The associated illness is typically mild and characterized by fever, headache, myalgia, and cutaneous manifestations (rash, eschar, and ulcer), but lymphadenopathy and neurological expressions (photophobia, hearing loss), abdominal symptoms (nausea, diarrhea), and pneumonia may also occur. Evidence of spotted fever group rickettsiosis was supported by serologic surveys and DNA analysis in the ROK (Lee et al. 1994, Jang et al. 2004, Choi et al. 2005), with human cases of infections reported in 2005 (Chung et al. 2006). In a previous study, we identified R. typhi in fleas captured in 2003–2004 (Kim et al. 2010b). While the fleas tested in the previous study had blood meals in their guts after removal from R. typhi–negative hosts, this does not preclude them from having taken recent blood meals from R. typhi–positive hosts.
A total of three species of fleas (C. congeneroides, R. insolita, and S. sidimi) collected from A. agrarius were found infected separately with R. typhi and R. felis. Doratopsylla coreana collected from C. lasiura was infected only with R. typhi. Similar to results by Noden et al. (1998), co-infections with both R. typhi and R. felis among individual fleas were not observed.
Human infections of murine typhus among Korean populations from 2005 to 2007 were more prevalent from October to November (46.1% and 36.7%, respectively), whereas for the other months, infection rates ranged from 0.0% to 4.1% (K-CDC 2009). Unfortunately, small mammal surveys were not conducted during October to determine flea indices for C. congeneroides, which was most frequently collected during August–September, and S. sidimi, which was very abundant during November–December (Kim et al. 2010b). Although it is not conclusive, data suggest that while S. sidimi appears to be the principal vector for R. typhi in the ROK, C. congeneroides, R. insolita, and D. coreana are also vectors. Data for spotted fever group Rickettsia, that is, R. felis, are not reportable events and therefore data for human cases are lacking.
Transmission of R. felis and R. typhi through fecal contamination of bite wounds or skin abrasions by these flea species that were PCR positive has not been verified. However, we detected R. felis and R. typhi DNA in spleen tissues of A. agrarius captured during a 2009 rodent survey using the same PCR method (data not shown). These results indicate that fleas that were positive for rickettsial parasites may have only contained parasites from recent blood meals taken from host animals. It is clear that the data on R. felis and R. typhi in A. agrarius are diverse in terms of the species implicated as hosts. Infections with R. felis and R. typhi have generally been considered to be host specific. Therefore, additional efforts to define the spectrum of host susceptibility in domestic and wild animals are needed. Although there are numerous reports throughout the world on the epidemiology of flea-borne pathogens and reservoir host/vector associations, this is the first report of R. felis from the ROK.
These studies provide further information on the epidemiology of flea-associated pathogens and the potential for transmission to humans in the ROK. Further studies are required for a better understanding of potential emerging zoonotic flea-borne pathogens in the ROK, which will enable health professionals to be more proactive regarding environmental changes and activities that impact the health of both civilian and military populations.
Footnotes
Acknowledgments
We thank the commanders and personnel of the 5th and 38th Medical Detachments, 168th Multifunctional Medical Battalion, for their support in conducting small mammal surveillance. We especially thank Dr. Joel Gaydos, Global Emerging Infections Surveillance and Response System, Silver Spring, MD, for his support and constructive criticism. Funding for portions of this work was provided by the Armed Forces Health Surveillance Center, Global Emerging Infections Surveillance and Response System, Silver Spring, MD, the National Center for Military Intelligence, Ft. Detrick, MD, and through the BK21 Program for Veterinary Science, Seoul National University, Seoul, Korea.
Disclosure Statement
The opinions expressed in this article are those of the authors and do not reflect official policy or positions of the U.S. Department of the Army, the U.S. Department of Defense, or the U.S. Government.