
Abstract
Canine babesiosis was considered an imported tick transmitted disease until the first case of autochthonous canine babesiosis in Slovakia was described in 2002. Since then, the number of cases kept increasing every year. The causative agent of babesiosis in dogs is not yet characterized; therefore, the aim of our study was to determine the agent and the rate of infection in the vector tick D. reticulatus in Slovakia. Babesia canis canis was detected in 80 out of 87 blood samples from dogs with clinical manifestations of babesiosis. Six dogs suspected of babesiosis tested positive for presence of Anaplasma phagocytophilum, and one mixed infection of B. c. canis and A. phagocytophilum was detected. B. c. canis was detected in 35.6% questing adults of D. reticulatus. The obtained sequences from blood samples showed 99.7% and from D. reticulatus, 99.4% similarity with the B. c. canis (AY072926) from dogs infected in Croatia. In our study, we characterized the agent of canine babesiosis from blood samples of naturally infected dogs and D. reticulatus, the vector tick. Further, the presence of A. phagocytophilum, bacterium responsible for the canine granulocytic anaplasmosis, was recorded in dogs for the first time in Slovakia.
Introduction
Occurrence of B. c. canis infections is limited to the presence of the tick vector D. reticulatus. In the last years, several European countries have noticed changes in the distribution of D. reticulatus tick, probably due to climate warming, landscape, and socio-economic changes (Bullova et al. 2009). However, the presence of B.c. canis was found also in Ixodes ricinus tick (Cieniuch et al. 2009). Clinical manifestation of canine babesiosis ranges from a hyperacute, shock-associated, hemolytic crisis to an inaparent, subclinical infection (Boozer and MacIntire 2003). Canine granulocytic anaplasmosis is another tick-transmitted disease that requires the attention of veterinarians in Central Europe. It is caused by gram-negative obligate intracellular bacterium Anaplasma phagocytophilum, which belongs to the order Rickettsiales, to the reorganized family Anaplasmataceae. A. phagocytophilum comprises three, previously single species E. phagocytophila, E. equi, and , now HGA (human granulocytic anaplasmosis) agent. Phylogenetic studies have revealed high nucleotide sequence similarity in their genomes, so they were unified into a single species A. phagocytophilum (Dumler et al. 2001). Anaplasma circulates in Europe between its main tick vector I. ricinus and various host animals (domestic and free-ranging). Besides canine and equine anaplasmosis, it causes HGA, tick-borne fever in sheep and pasture fever in cattle (Rikihisa 1999). Disease caused by A. phagocytophilum appears to be less severe than in cases of that caused by E. canis, E. ewingii, or B. c. canis (Preziosi and Cohn 2002, McQuiston et al. 2003).
The aim of our study was to identify and characterize the Babesia and other possible tick-transmitted pathogens in the blood of dogs with clinical signs of babesiosis and the prevalence of Babesia in the vector tick D. reticulatus in the area with endemic occurrence of canine babesiosis in Slovakia.
Materials and Methods
Blood samples and tick collection
During the outbreak of babesiosis in the years 2004–2005 and till the year 2010, veterinary practitioners from areas with endemic occurrence of canine babesiosis in the southern part of Slovakia were asked to collect blood samples from dogs showing clinical signs of babesiosis. Blood samples were collected into EDTA (8% liquid solution) tubes and kept at +4°C until the DNA isolation.
In the same area, the questing adults of D. reticulatus ticks were collected by flagging the vegetation. Localities with increased presence of dogs were chosen, especially the central parks near housing estates and the banks of rivers flowing directly through them. Ticks were immediately stored in 70% ethanol.
DNA isolation
DNA was isolated from 200 μL of EDTA-anticoagulated blood of each dog using a QIAamp DNA blood mini kit (Qiagen, Hilden, Germany). Extraction steps were carried out according to the manufacturer's protocol.
DNA from individual ticks was isolated using DNeasy tissue kit (Qiagen, Hilden, Germany). Before extraction, the ticks were immediately dried for 30 min to evaporate the ethanol. Each tick was cut with a disposable sterile scalpel. The steps described above were carried out according to the manufacturer's protocol. DNA was stored in +4°C.
Polymerase chain reaction (PCR)
PCR amplification was performed in a total of 25-μL reaction mixture MasterTaq DNA polymerase kit (Eppendorf AG, Hamburg, Germany) containing 10.4 μL of deionized water, 5 μL of 5× TaqMaster PCR Enhancer, 2.5 μL of 10× Taq buffer (with 15 mM Mg2+), 1.5 μL of a 25 mM solution of Mg (OAc) 2, 0.1 μL of Taq DNA polymerase (5U/μL), 0.5 μL of dNTP-mix (10 mM) (Fermentas, Vilnius, Lithuania), 1.25 μL of each primer (10pmole/μL) (Invitrogen, Paisley, Scotland), and 2.5 μL of DNA template. In this study, two sets of primers for Babesia were used: Primer Babesia-F (5′-GTG-AAA-CTG-CGA-ATG-GCT-CA-3′) and primer Babesia-R (5′-CCA-TGC-TGA-AGT-ATT-CAA-GAC-3′), which were designed on the basis of the common sequence of the 18S rRNA gene of the genus Babesia; the PCR product has an approximately 650-bp (Inokuma et al. 2003) and was used for the detection of babesiae in blood sampes. The amplification procedure was performed as previously reported (Inokuma et al. 2000). Tick DNA samples were examined for the presence of B. c. canis using the pair of primers 793-772R (5′-ATG-CCC-CCA-ACC-GTT-CCT-ATT-A-3′) and BCC-F (5′-TGC-GTT-GAC-GGT-TTG-ACC-3′) designed for the B. c. canis specific PCR based on the 18S rRNA gene, which amplify 198-bp amplicons (Birkenheuer et al. 2003). The primers EHR 521 (5′-TGT-AGG-CGG-TTC-GGT-AAG-TTA-AAG-3′) and EHR 747 (5′-GCA-CTC-ATC-GTT-TAC-AGC-GTG-3′) were used to amplify an approximately 250 bp region of the 16S rRNA gene of the family Anaplasmataceae in the DNA (Pancholi et al. 1995). Moreover, to confirm the presence of A. phagocytophilum and to exclude detection of other members of the Anaplasmataceae, we used nested PCR specific for the detection of A. phagocytophilum using primers amplifying a fragment of 16S rRNA, ge3a (5′-CAC-ATG-CAA-GTC-GAA-CGG-ATT-ATT-C-3′), and ge10r (5′-TTC-CGT-TAA-GAA-GGA-TCT-AAT-CTC-C-3′) for the first-round amplification. One microliter of the first-round PCR product was used as a template for the second PCR with the set of primers: ge9f (5′-AAC-GGA-TTA-TTC-TTT-ATA-GCT-TGC-T-3′) and ge2 (5′-GGC-AGT-ATT-AAA-AGC-AGC-TCC-AGG-3′). PCR conditions were used according to (Massung et al. 1998). The PCR products were electrophoresed on a 1% agarose gel, stained with ethidium bromide, and visualized with a UV transilluminator.
DNA sequencing of PCR products
Sequencing was performed at the Danube Clone Ltd. Sequencing facility at the Institute of Neuroimmunology (Slovak Academy of Sciences, Bratislava, Slovak Republic). Randomly selected PCR products of Babesia from blood and ticks and all PCR products of ehr521-ehr747 primers were sequenced. PCR products were purified by using a QIAquick PCR purification kit (Qiagen). The complementary strands of each sequenced product were manually assembled. Sequences were compared with GenBank entries by Blast N2.2.13. Homologous sequences were aligned using the CLUSTAL W Multiple Sequence Alignment Program (version 1.81). Sequence similarities among the sequences were calculated by EMBOSS Align, a pairwise alignment algorithm (
Results
Clinical manifestation
Detailed clinical signs were recorded in 35 cases. Nonspecific signs such as weakness, apathy, loss of appetite, increased heart rate, and breath frequency were recorded in each case. Body temperature ranged from 38.9°C–40.9°C. Specific signs with different frequency were observed (Table 1).
Examination of blood samples
A total of 87 blood samples with clinical manifestation of babesiosis were obtained from veterinary practitioners. All samples were analyzed for the presence of Babesia. The fragment of 18S rRNA gene of the genus Babesia was amplified using Babesia F-Babesia R primers in samples. Three randomly chosen samples were sequenced. The obtained sequences (accession number: DQ869307) showed 100% homology to one another and 99.7% similarity with the B. c. canis (AY072926) from dogs infected in Croatia (Caccio et al. 2002). According to the classification of Adaszek and Winiarczyk (2008), our strains of B. c. canis belong to the group B, they have AG in the position 150 and 151 of 18S rRNA. Moreover, all positive samples were tested using B. c. canis specific primers to confirm the presence of the agent. Its presence was confirmed in all 80 samples collected in southern Slovakia (Fig. 1).
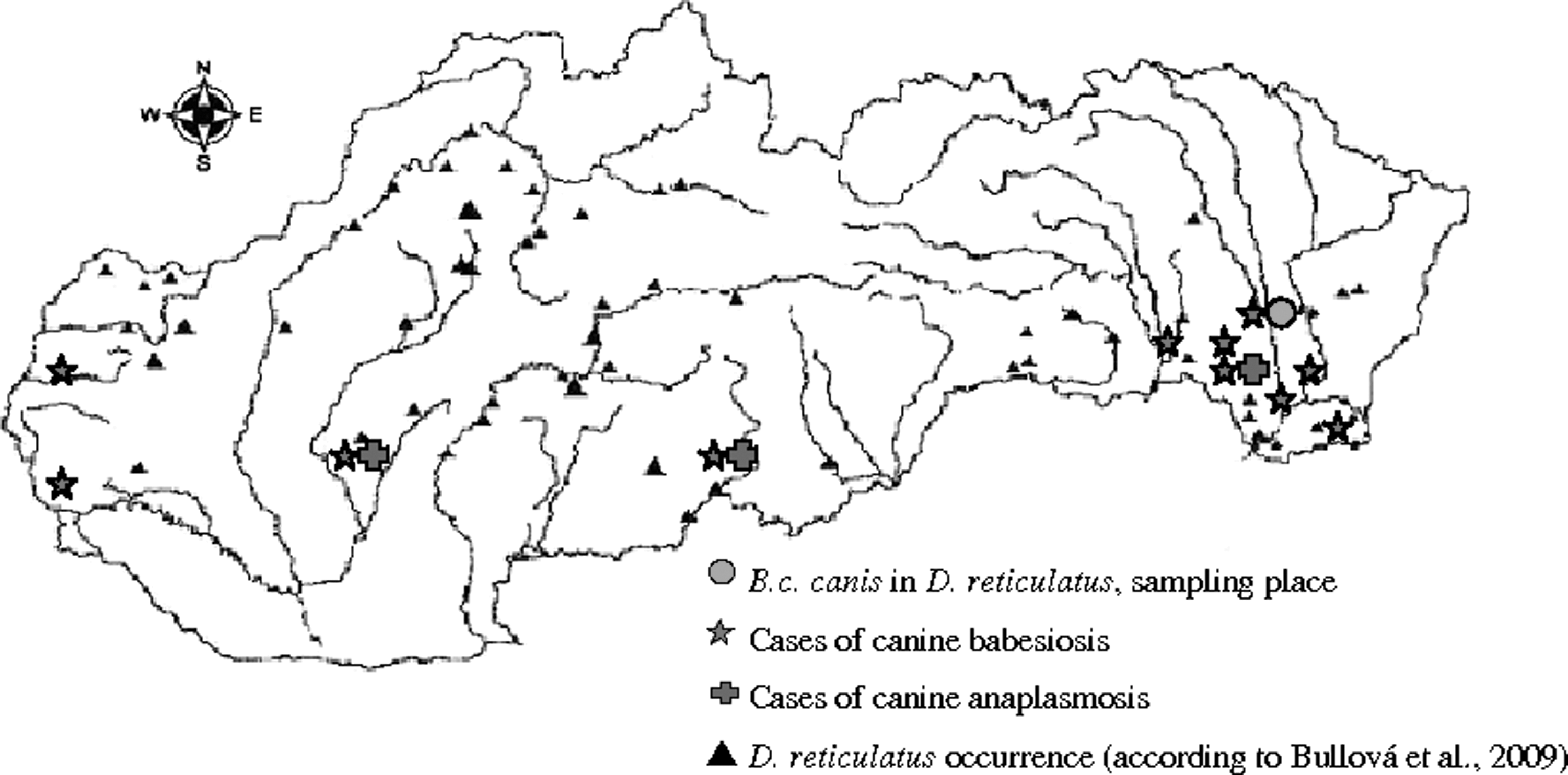
Occurrence of cases of canine babesiosis and anaplasmosis in Slovakia. The distribution of D. reticulatus according to Bullová et al., 2009. Largest rivers outlined.
All 87 blood samples were tested with primers detecting members of the family Anaplasmataceae. Seven samples tested positive. Two samples were sequenced, and the obtained sequences DQ907007 and GU724971 showed 100% similarity to A. phagocytophilum strains isolated from rodents (HM439430), cervids (AF481854), and vector ticks (HQ629925). Moreover, to confirm the presence of A. phagocytophilum and to exclude detection of other members of the Anaplasmataceae, we used nested PCR specific for the detection of A. phagocytophilum. In all seven samples, PCR product was detected and A. phagocytophilum was confirmed.
One mixed infection of B. c. canis and A. phagocytophilum was detected in a dog with clinical signs of babesiosis.
Examination of ticks
A total of 326 (204 females and 122 males) questing adult D. reticulatus ticks were collected by flagging (Fig. 1). PCR using B. c. canis specific primers revealed positive results in 116 (35.6%) ticks. The prevalence in female (36.3%) ticks was higher than in males (34.4%). The difference was not significant. Five randomly selected positive samples were reamplified using Babesia F-Babesia R primers to obtain a longer PCR product for further sequencing. All five obtained sequences were identical (accession number DQ869308) and showed 99.4% similarity with sequences of Babesia from canine blood samples from that area, and 99.1% similarity to the B. c. canis (AY072926) from an infected dog in Croatia, and belong to the B group of B. c. canis.
Discussion
In Slovakia, canine babesiosis was considered an imported tick-transmitted disease until the first case of autochthonous infection in dog was described in 2002 by Chandoga et al. (2002). Since then, the number of cases has been increasing every year. The causative agent of canine babesiosis in Slovakia has not yet been characterized at the molecular level; therefore, the aim of our study was to identify the agent and the rate of infection in the vector tick D. reticulatus as well as in the dogs with clinical signs. Canine babesiosis is highly endemic in France, Switzerland, Hungary, Serbia, Croatia, northern Italy, and northern Spain but it was traditionally less common in Belgium, Germany, Poland, and The Netherlands (Beck et al. 2009, Beugnet and Marie 2009). Babesiosis has been previously reported in Hungary, bordering Slovakia on the South (Horváth and Papp 1996). The distribution of the tick vector together with the occurrence of the disease is spreading to new regions (Foldvari et al. 2005, 2007).
Light microscopic examination of stained blood smears is the standard diagnostic protocol for the veterinarians. In April 2004, during the outbreak of canine babesiosis, 20 cases per day with clinical signs of this disease were recorded in a small animal practice in South-eastern Slovakia. The activity of D. reticulatus, the vector tick and the reservoir host of this piroplasm, usually peaks during this month.
B. c. canis subspecies specific PCR were used to confirm the causative agent of babesiosis. The primers were designed, and the specificity was tested by Birkenheuer et al. (2003) on the DNA isolated from B. c. canis. B. c. rossi, B. c. vogeli, B. gibsoni, and T. annae-infected canine whole-blood samples. Results confirmed that the pair of primers 793-772R and BCC-F is specific for B. c. canis.
In relation to environmental changes, new localities suitable for tick survival are emerging in the areas where conditions were not favorable in the past. This fact also affects the epidemiology of tick-borne diseases and formation of new host-parasite relations. These changes support expansion of the areas of occurrence and emergence of new pathogenic organisms in Slovakia. D. reticulatus was known to occur in western Slovakia (Malacky and along the Danube River) and in parts of eastern Slovakia (along the Latorica River) (Nosek 1972). In the survey of Bullova et al., 2009 new areas with D. reticulatus occurrence were detected, providing evidence that this tick species has extended its range not only in the surroundings of its former habitats but also by at least 200 km further north and by 300 m of elevation into higher altitudes in Slovakia.
Similarly, Rar et al. (2005) reported that the distribution of the vector D. reticulatus is changing, the tick is spreading its range of occurrence. Siuda et al. (1998) found that D. reticulatus is slowly extending its range from the east to the west of Poland. Vector competence of D. reticulatus for B. c. canis is now indisputable. We detected 35.6% prevalence of B. c. canis in D. reticulatus. Our findings are in contrast to 1% prevalence found by (Duh et al. 2006) in Dermacentor ticks collected in western Slovakia as well as to 4.16% prevalence detected by Rar et al. (2005). D. reticulatus is present in England and Wales (Hillyard 1996), but the cases of autochthonous canine babesiosis have never been reported there. Accessibility and high abundance of competent hosts, high ratio of transovarial transmission, geographical, ecological, and other circumstances can increase the infection rate of the pathogen in the vector ticks. High prevalence of Babesia in ticks in our study suggests a possible relation to increased dog presence in localities where the ticks were flagged in contrast to low prevalence in ticks collected in the forest (Duh et al. 2006).
Canine anaplasmosis with similar clinical manifestation as babesiosis in the early stage can be misdiagnosed as babesiosis, and the real cause of the disease is hidden. In our study, we determined 6 cases of canine anaplasmosis and 1 mixed infection of A. phagocytophilum and B. c. canis. Diagnosis of canine babesiosis cannot be based on the observed clinical signs only (Foldvari et al. 2005), and differential diagnostics should be emphasized. The finding of A. phagocytophilum in dogs with clinical signs represents the first case of infection of the dog in Slovakia.
Currently, due to various factors that are linked to the changing epidemiology of vector-borne disease emerging and re-emerging, diseases represent a risk for dogs. Dirofilariosis appears to have remained epidemiologically stable in most of Europe (Miterpakova et al. 2008, Duscher et al. 2009, Jacso et al. 2009). Hepatozoonosis is extending the range of occurrence (Vojta et al. 2009, Dezdek et al. 2010), and agents of canine leishmaniosis and monocytic ehrlichiosis are spreading toward north (Beugnet and Marie 2009).
In our study, we characterized the agent of canine babesiosis from blood samples of naturally infected dogs as well as in D. reticulatus, the vector tick. Moreover, in seven dogs suspected of babesiosis, A. phagocytophilum was detected, suggesting that canine anaplasmosis may be often undetected or misdiagnosed and left untreated. Therefore, veterinary practitioners should particularly emphasize on differential diagnostics to initiate appropriate therapy. In areas where canine babesiosis is endemic, pet owners should pay attention to tick prevention and control.
Footnotes
Acknowledgments
The publication has been realized within a frame of the project Centre of Excellence for Parasitology (Code ITMS: 26220120022) based on the support of the Operational Programme “Research & Development” funded from the European Regional Development Fund (rate 0.2), by the Slovak Agency of Research and Development, APVV 0108-06 and LPP 0341-06, by the Scientific Grant Agency of the Ministry of Education of Slovak Republic, and by the Slovak Academy of Sciences no 2/0128/09.
Disclosure Statement
No competing financial interests exist.