
Abstract
In some rodent species frequently exposed to plague outbreaks caused by Yersinia pestis, resistance to the disease has evolved as a population trait. As a first step in determining if plague resistance has developed in black-tailed prairie dogs (Cynomys ludovicianus), animals captured from colonies in a plague-free region (South Dakota) and two plague-endemic regions (Colorado and Texas) were challenged with Y. pestis at one of three doses (2.5, 250, or 2500 mouse LD50s). South Dakota prairie dogs were far more susceptible to plague than Colorado and Texas prairie dogs (p<0.001), with a mortality rate of nearly 100% over all doses. Colorado and Texas prairie dogs were quite similar in their response, with overall survival rates of 50% and 60%, respectively. Prairie dogs from these states were heterogenous in their response, with some animals dying at the lowest dose (37% and 20%, respectively) and some surviving even at the highest dose (29% and 40%, respectively). Microsatellite analysis revealed that all three groups were distinct genetically, but further studies are needed to establish a genetic basis for the observed differences in plague resistance.
Introduction
In North America, prairie dogs (Cynomys spp.) have long been considered epizootic hosts for plague and highly susceptible to the disease (Cully and Williams 2001). Plague was first confirmed in black-tailed prairie dogs (BTPDs; Cynomys ludovicianus) in 1947 in Kansas (Cully et al. 2000) and Texas (Miles et al. 1952), and even earlier (1936–38) in Utah prairie dogs (Cynomys parvidens; Utah), white-tailed prairie dogs (Cynomys leucurus; Wyoming) and Gunnison's prairie dogs (Cynomys gunnisoni; New Mexico; Eskey and Haas 1940). In the early 1950s, the eastern boundary of plague extended to about the 103rd meridian (Fig. 1) and remained there with little deviation for over 50 years (Adjemian et al. 2007). Within the plague-endemic states, mortality in BTPDs often reaches nearly 100% during plague epizootics (Cully and Williams 2001), but presumed survivors have been observed after outbreaks (Pauli et al. 2006), and a few animals nearly always survive experimental infection, even after injection of high numbers of bacteria (Mencher et al. 2004, Rocke et al. 2008, 2010). The extreme sociality of prairie dogs in relation to other rodent species probably contributes significantly to their susceptibility to a vector-borne disease like plague (Hoogland 1995). Biggins and Kosoy (2001) suggested that the costs of the prairie dogs' social system may outweigh benefits in the long term unless prairie dogs become more resistant to plague, as have other highly social rodents in plague-endemic foci in Asia. Given the extreme selective pressure of plague on prairie dogs, it is reasonable to hypothesize that some degree of natural resistance to the disease would eventually evolve in populations with a history of exposure, although to date, no direct evidence of resistance in any prairie dog species exists. Using BTPDs captured in 2003 (Fig. 1) from Colorado and Texas (considered plague-endemic states) and South Dakota (a plague-free area at that time), we tested the hypothesis that susceptibility to experimental plague infection would differ between populations from plague-endemic and plague-free regions. Microsatellite analyses of DNA samples were used to verify population differences at neutral genetic loci among the three populations.
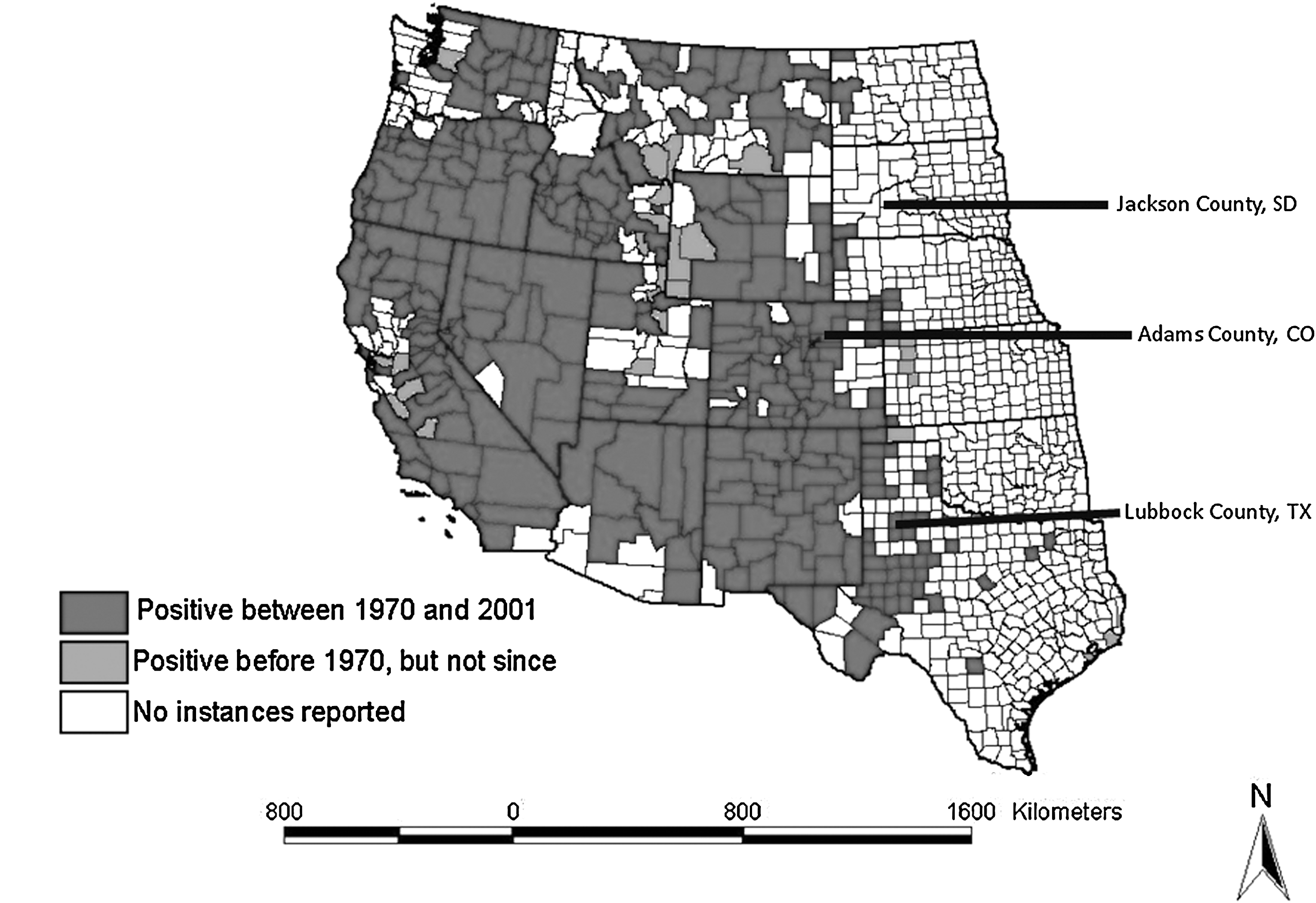
Prairie dogs capture sites in relation to plague occurrence in the United States (based on Antolin et al. 2002).
Materials and Methods
Experimental animals
Free-ranging adult BTPDs captured in 2003 from colonies in South Dakota (Jackson County; n=24), Colorado (Adams County; n=26), and Texas (Lubbock County; n=17) were transported to the U.S. Geological Survey National Wildlife Health Center (NWHC, Madison, WI). Specific trapping locations are listed in Table 1. At the Colorado and Texas sites, significant outbreaks of plague had not been observed before our study for at least 2 and 10 years, respectively, according to local public health officials. Before shipment, the animals were dusted with carbaryl, and, upon arrival at NWHC, they were inspected for external parasites (none were found), injected with an anthelminthic (200 μg/kg Ivermectin; Merck & Co., Inc), and treated with 200 μL of Advantage flea control (Imidacloprid; Bayer HealthCare, Animal Health Division, Shawnee Mission, KS) via external application to the skin on the back of the neck. Electronic microchip identification units (Avid Identification Systems, Inc.) were inserted into each animal between the scapulae via subcutaneous injection. Prairie dogs were group-housed by population in three isolation rooms with ∼180 sq ft of floor space. Bedding material (Beta chips) covered the floor and custom-made stainless steel nest boxes with connecting lengths of polyvinyl chloride pipe were used for shelter. Alfalfa-based pellets (∼50 g per animal per day) and fresh vegetables (broccoli, carrots, green beans, and sweet potato chunks) were provided once daily. Water was available ad libitum.
Y. pestis challenge
Prairie dogs were acclimated at NWHC for at least 6 weeks before challenge. In each room, prairie dogs were randomly assigned to receive a low, medium, or high challenge dose of Y. pestis in roughly equivalent numbers (Table 1). Two animals in each room were not inoculated but served as contact controls to determine if additional transmission between animals occurred. All prairie dogs were inoculated on the same day with dilutions of the same inocula. To prepare the inocula, a stock aliquot of the bacteria (the CO92 wild-type isolate of Y. pestis provided by the U.S. Army Medical Research Institute of Infectious Diseases), prepared and quantified as previously described (Osorio et al. 2003), was diluted in phosphate-buffered saline to provide 50, 5000, and 50,000 cfu, which corresponds to 2.5, 250, and 2500 mouse LD50, respectively. A volume of 0.2 mL of this solution was administered to each prairie dog by subcutaneous injection in the right hip region. Plate counts of the challenge inocula confirmed the dosages and concurrent mouse tests confirmed their expected virulence. Prairie dogs were monitored for 28 days for signs of illness or death. Animals with obvious clinical signs (labored breathing, disinclination to move) were humanely euthanized as were all survivors at the end of the 28-day period. Carcasses were frozen for future necropsy.
Plague-induced mortality was verified by isolation of Y. pestis–specific DNA sequences from tissue culture by polymerase chain reaction (PCR) assays. Frozen carcasses were thawed and necropsied. Skin samples were collected from every individual and frozen for later DNA analysis. Tissue samples from lung, liver, and spleen were collected from selected animals, and these were cultured in brain heart infusion broth (Difco) and on blood agar plates (Becton-Dickinson) at 28°C for up to 72 h. DNA was subsequently extracted from the culture broth and stored at −20°C. For PCR, primers specific for the Y. pestis F1 gene (Heath et al. 1998) were used to amplify DNA fragments that were fractionated and directly observed using standard agarose gel techniques.
Serology
Blood samples (300 μL) were collected from the medial saphenous vein of each prairie dog before challenge and postchallenge from all those that survived infection. Sera were collected and stored at −20°C until analyses. Antibody titers to Y. pestis F1 and V antigens were determined using enzyme-linked immunosorbent assays with antigens supplied by the U.S. Army Medical Research Institute of Infectious Diseases as described previously (Rocke et al. 2008). Briefly, serum samples were serially diluted fourfold from 1:160 to 1:40,960; test samples were run in duplicate. Samples with titers <1:160 were recorded as 1:40. Each plate also contained four replicates of a negative control serum sample and two replicates of a positive control serum sample. A horseradish-peroxidase-labeled anti-prairie dog IgG custom-prepared by Bethyl Laboratories was diluted 1:100 and used as the secondary antibody. Titers <1:160 were considered negative. The highest dilution that was positive (exceeded the mean of four negative control samples by three standard deviations) was considered the endpoint and its reciprocal value recorded as the titer.
Genetic analyses
Skin samples were processed by scraping away the hair and removing a 20 mg portion for DNA extraction using the Wizard SV Genomic DNA Purification System (Promega). To determine genetic differentiation among the prairie dog samples from these populations, we generated multilocus genotypes for all individual prairie dog samples using a set of 11 neutral microsatellite markers originally developed for BTPDs and ground squirrels (Spermophilus spp.) (May et al. 1997, Stevens et al. 1997, Roach et al. 2001, Jones et al. 2005). These microsatellite markers were multiplexed according to annealing temperature, except marker D2 was run alone. The PCRs for nine of the markers were carried out in 10 μL reactions containing 40 ng of genomic DNA, 1 μL 10×PCR buffer, 2 mM MgCl2, 0.2 mM dNTPs, 1 U Platinum Taq polymerase (Invitrogen), and 0.1–0.4 μM of each primer. The PCR profile for these nine markers started with an antibody-release step at 95°C for 10 min, followed by 34 cycles of 1 min at 94°C, 30 s of annealing, and 30 s at 72°C, with 5 min of final extension at 72°C. The PCRs for the remaining two markers (GS-14 and GS-22) contained 1.5 mM MgCl2, 0.12 mM dNTPs, and 0.2 μM of each primer and the PCR profile followed that of Stevens et al. (1997), except for a 95°C hot start for 10 min and an annealing temperature of 53°C during the 33 cycles. PCR products were genotyped using an ABI 3130 sequencer and analyzed using the software GENEMAPPER v4.0 (Applied Biosystems).
Statistical analyses
Survival data were analyzed with the Cox proportional hazards model (Cox 1972). A two-way design with interaction (population by challenge dose) and all nested sub-models were fitted with SAS PROC PHREG (SAS Institute Inc.). Kaplan–Meier survival curves were also calculated using SAS.
We assessed genetic divergence among the three BTPD populations using F ST and assignment approaches. Global and pairwise F ST values were estimated using the program SPAGeDi (Hardy and Vekemans 2002), whereas PhiST was estimated using an AMOVA in GENALEX (Peakall and Smouse 2006). We also used assignment testing to check for any admixture in these gene pools using STRUCTURE (Pritchard et al. 2000). Bayesian priors were set to assume no admixture and independent allele frequencies, which is appropriate given the large geographic distances separating each collection site. We used a burn-in time of 25,000 iterations followed by data collection over an additional 100,000 iterations. Finally, we used GENALEX to compare mean pair-wise relatedness (Queller and Goodnight 1989) of survivors versus susceptible animals for each population separately. This allowed us to check for any obvious patterns due to related individuals.
Results
The numbers of animals that survived plague challenge at each dose within each group are listed in Table 1. Plague mortality was highest (nearly 100%) in animals captured at the South Dakota site, regardless of dose. In contrast, mortality was much lower in animals captured at the Colorado and Texas sites at all doses. When the interaction model was compared with the additive two-way model using the Cox model, there was no evidence for a dose by capture site interaction (G2=0.7, df=4, p>0.5). When the additive two-way (dose and site) model was compared to the site-only model, there was only weak evidence for a dose effect, which did not achieve statistical significance (G2=4.1, df=2, p=0.129). When the additive two-way model was compared to the dose-only model, there was strong evidence for a site effect (G2=21.5, df=2, p<0.001). As evidenced by the hazard ratios (Table 2) and Kaplan–Meier survival curves (Fig. 2), the survival rates of prairie dogs captured at the Colorado and Texas sites were substantially higher than animals from the South Dakota site (Table 2; Fig. 2). No difference was detected in survival rates between Colorado and Texas prairie dogs. Y. pestis was confirmed from all carcasses tested. None of the uninfected contact controls became sick or died in any of the groups indicating further transmission of the bacteria among individuals within groups did not likely occur.
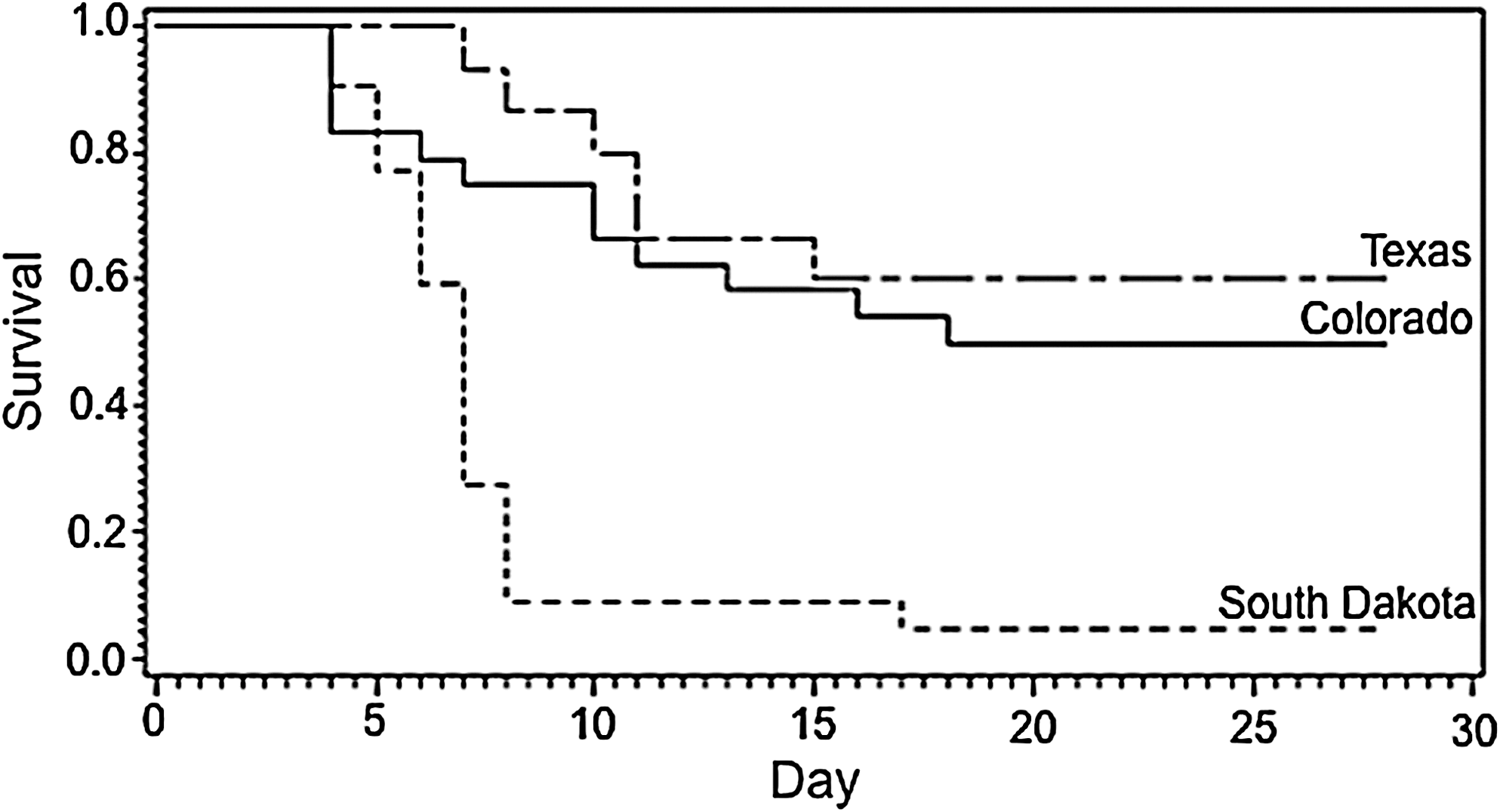
Kaplan–Meier survival curves of black-tailed prairie dogs captured at different locations in Colorado (CO), South Dakota (SD), and Texas (TX) and challenged with Yersinia pestis.
Serology
No significant antibodies to F1 and V antigens were detected in samples collected from prairie dogs prechallenge. Only 5 of the 21 animals that survived infection developed significant anti-F1 antibody titers (>1:160) postchallenge and even fewer (two) had antibody titers to V antigen.
Genotyping
The 11 microsatellite markers displayed a considerable amount of allelic diversity across these populations and observed heterozygosity was fairly high (Table 3). None of the 59 samples had identical multilocus genotypes. The F ST estimate was 0.17 (p<0.001), indicating a large amount of genetic differentiation among populations. All pairwise comparisons displayed about the same level of divergence, with pairwise F ST estimates ranging from 0.15 to 0.17. The AMOVA estimate also reflected this high level of genetic structure (PhiST=0.16, p=0.001). The STRUCTURE analysis was most consistent with three independent gene pools with no gene flow between them. These results are congruent with the wide geographic spread of our three prairie dog populations and with previous population genetic analyses of BTPDs (Roach et al. 2001, Antolin et al. 2006). Pair-wise relatedness values of susceptible versus resistant prairie dogs within groups were all below zero, indicating relatedness was not a factor in immunity to plague (mean r xy values: CO susceptible=−0.055, CO resistant=−0.062; SD susceptible=−0.053, SD resistant=no pairwise comparisons available; TX susceptible=−0.093, TX resistant=−0.123).
Stevens et al. (1997).
May et al. (1997).
Jones et al. (2005).
T a, annealing temperature (°C); H O, observed heterozygosity.
Discussion
The results of our study provide the first evidence that BTPDs in plague-endemic regions of the United States may have evolved some resistance to Y. pestis. Animals captured in plague-endemic sites (Colorado and Texas) were considerably more resistant to laboratory challenge with Y. pestis than BTPDs captured from a plague-free site (South Dakota), regardless of challenge dose. Further, prairie dogs from the plague-endemic sites were heterogenous in their response, with some animals dying at the lowest dose (37% and 20%, respectively) and some surviving even at the highest dose (29% and 40%, respectively).
Microsatellite analysis confirmed that these three groups of BTPDs were genetically distinct at neutral markers, despite similarities in resistance in animals from the plague-endemic sites (Texas and Colorado). Additional work is needed to confirm that differences in plague susceptibility between sites were genetically based. Other nongenetic explanations for the observed differences, such as physiologic condition and immune status, are possible but unlikely. Antibodies indicative of prior exposure to plague were not detected in any of the animals before the laboratory challenge, and evidence of a booster response in survivors postchallenge was likewise absent. The lack of significant levels of postchallenge antibodies is consistent with our previous vaccine and challenge studies (Rocke et al. 2008, 2010), indicating that innate or nonspecific mechanisms may be more important to surviving plague infection than adaptive immunity, at least for prairie dogs. This premise is consistent with Y. pestis virulence mechanisms, which are known to predominantly target the innate immune system (Kerschen et al. 2004, Bergsbaken and Cookson 2009). Infection with other agents that might provide cross-protection against plague is possible. However, the agents most likely to cross-protect are other Yersinia spp. that share the crV gene in common with Y. pestis, and no anti-V antibody was detected in animals before challenge. We hypothesize that the resistance we observed in animals from the plague-endemic sites is genetically based, has evolved independently multiple times through natural selection, and is dependent on innate immune mechanisms.
Because prairie dog colonies (of all species) suffer extremely high rates of mortality during plague epizootics and even local extinctions due to the disease, the evolution of plague resistance in prairie dogs was previously thought unlikely or nonexistent (Cully et al. 2000, Roach et al. 2001, Gage and Kosoy 2005). Higher plague resistance in rodent populations from plague-endemic areas has been demonstrated in other North American wild rodents, including voles (M. californicus; Hubbert and Goldenberg 1970), grasshopper mice (Thomas et al. 1988), and California ground squirrels (Spermophilus beecheyi; Meyer 1942). These species have been considered enzootic or maintenance hosts for the disease with lower mortality rates than prairie dogs and, thus, a greater capability to develop even higher rates of resistance. Prairie dogs have typically been thought of as epizootic or secondary hosts that cannot maintain the disease between epizootic periods because of their extreme susceptibility to the disease (Biggins and Kosoy 2001). However, results of recent field studies using treatments that attempt to control for plague on half the study site (through plague vaccination and pesticide application) challenge this notion and suggest that plague may occur at very low or enzootic levels in at least some BTPD colonies (Biggins et al. 2010, Matchett et al. 2010). Because we do not know how often prairie dog colonies may be exposed to plague, it is impossible to assess if the frequency of epizootics in specific populations has stayed the same, increased, or decreased over time, possibly in response to evolving resistance, or if the frequency of enzootic plague in prairie dogs may be increasing. Plague epizootics with high mortality rates obviously still occur in BPTDs in the plague-endemic region, but perhaps higher levels of bacteria, greater flea burdens, or some other key factors are required to initiate epizootics in host populations that are becoming more resistant to the disease.
Controlled experimental infection studies, such as ours, are the best way to assess and compare plague resistance among populations within a species. To follow-up our studies and confirm the genetic basis of plague resistance in BTPDs, studies on F1 progeny from each of the selected populations and between population hybrids (e.g., Colorado and South Dakota animals), bred and maintained under the same conditions, would be very useful, much like studies in grasshopper mice (Thomas et al. 1988) and mid-day gerbils (Meriones meridianus; Levi 1994). Genomic analyses of F1 progeny and hybrids in relation to disease outcome would allow exploration and identification of genetic markers associated with plague susceptibility in this species. Recent studies in Madagascar black rats identified several amplified fragment length polymorphism markers associated with a plague-resistant phenotype from plague-endemic regions as well as resistance to plague challenge in experimental infection studies (Tollenaere et al. 2011), an important first step in identifying the underlying gene or genes involved in plague resistance. Analyzing plague resistance in BTPDs and other prairie dog species would provide a similar opportunity to evaluate pathogen-mediated natural selection to an invasive disease in a highly susceptible host.
Footnotes
Acknowledgments
This work was funded by the U.S. Geological Survey and NIH-NIAID via the Pacific-Southwest RCE (AI065359). The authors are grateful to P. Nol and S. Smith for technical assistance; D. Heisey for statistical advice; and D. Biggins, J. Hall, and R. van Andel for editorial comments. Use of trade, product, or firm names does not imply endorsement by the U.S. Government.
Disclosure Statement
No competing financial interests exist.