
Abstract
West Nile virus (WNV) ecology in neotropical ecosystems is poorly understood, and vertebrate hosts responsible for infecting mosquitoes remain unidentified throughout the Caribbean Basin. After a period of intense WNV transmission among sentinel chickens near Ceiba, Puerto Rico, we measured abundance of resident birds and species-specific prevalence of WNV infection. Taking the product of these measures indicates the relative number of WNV infections by species. Greater Antillean grackle (Quiscalus niger) accounted for the most WNV infections among birds in our 100-km2 study site. In urban habitats, the house sparrow (Passer domesticus) was frequently infected. Immature birds less than one year of age were more likely to have detectable WNV-reactive antibodies than older birds of the same species.
Introduction
The lack of WNV disease outbreaks in subtropical and tropical regions despite the presence of transmission remains enigmatic. One theory suggests that hyperendemic dengue virus transmission stimulates cross-protective anti-flavivirus immunity in people, but this theory fails to explain protection in horses and birds (Bosch et al. 2007). In the United States, certain bird species drive the WNV transmission cycle (Komar 2003). However, WNV transmission dynamics in the tropics may differ from temperate ecosystems, such that birds play a lesser role as amplifying hosts, or WNV genotypes circulating in the tropics may exhibit reduced virulence relative to those in temperate North America. The principal amplifying hosts, that is, the vertebrate species primarily responsible for infecting mosquitoes with WNV, have not yet been characterized anywhere in the Caribbean Basin.
Low-level WNV transmission was first detected in Puerto Rican birds and horses in 2004 near Ceiba (Dupuis et al. 2005, Barrera et al. 2008,). To study WNV ecology there, a network of sentinel chickens was established in July 2006 (Barrera et al. 2008). Presumably, certain common species of resident passerine birds drive transmission in Puerto Rico. Accordingly, we estimated the relative abundance and prevalence of WNV infection in free-ranging bird species resident near Ceiba shortly after a period of intense transmission had occurred in sentinel chickens in 2007. These data were then used to infer the relative number of avian WNV infections by species within our study site.
Methods
Study area
The region selected for study is approximately 100 km2 and encompasses the Roosevelt Roads Naval Base and parts of Naguabo and Ceiba municipalities in eastern Puerto Rico (Figs. 1 and 2). Selection criteria included previous detection of WNV-infected birds and horses here in 2004, presence of multiple habitats, and evidence of recent WNV transmission among sentinel chicken flocks in 2007 (Dupuis et al. 2005, Barrera et al. 2008). Ikonos satellite images of the site were visually examined to estimate the area of each principal habitat (Table 1). Habitats included upland forest (including humid evergreen forest and dry semideciduous forest), mangrove forest (includes associated tidal flats), open land (mowed fields and lawns, airport runways, and parking lots), rural (cropland, pasture, marshes, managed landscapes, and low-density residential neighborhoods), urban (high-density residential neighborhoods, and commercial districts), and water (lakes, lagoons, and bays).
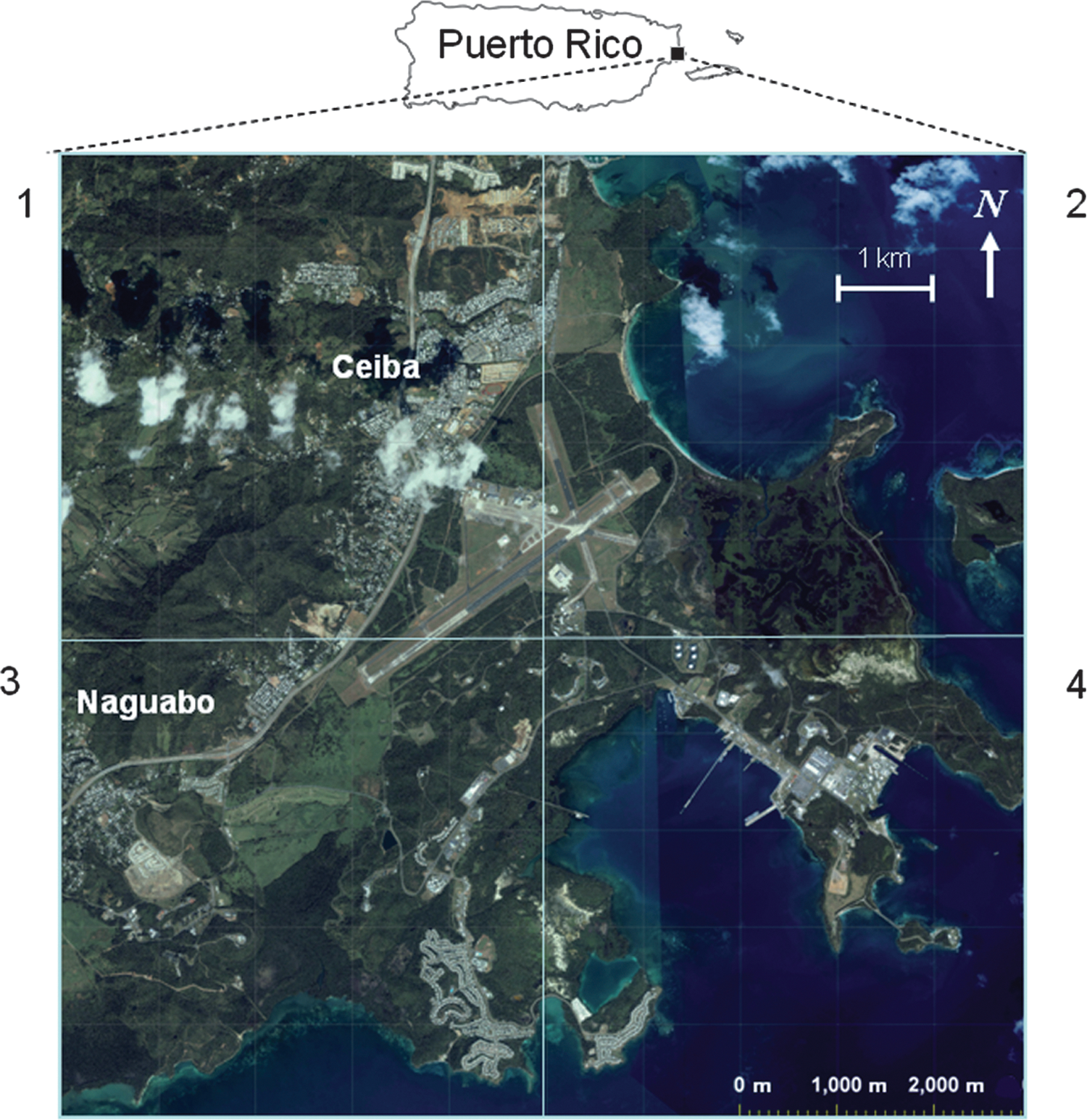
Aerial photograph of 100-km2 study area in the vicinity of Ceiba, Puerto Rico, indicating four quadrants used to estimate relative area of various habitat classes.
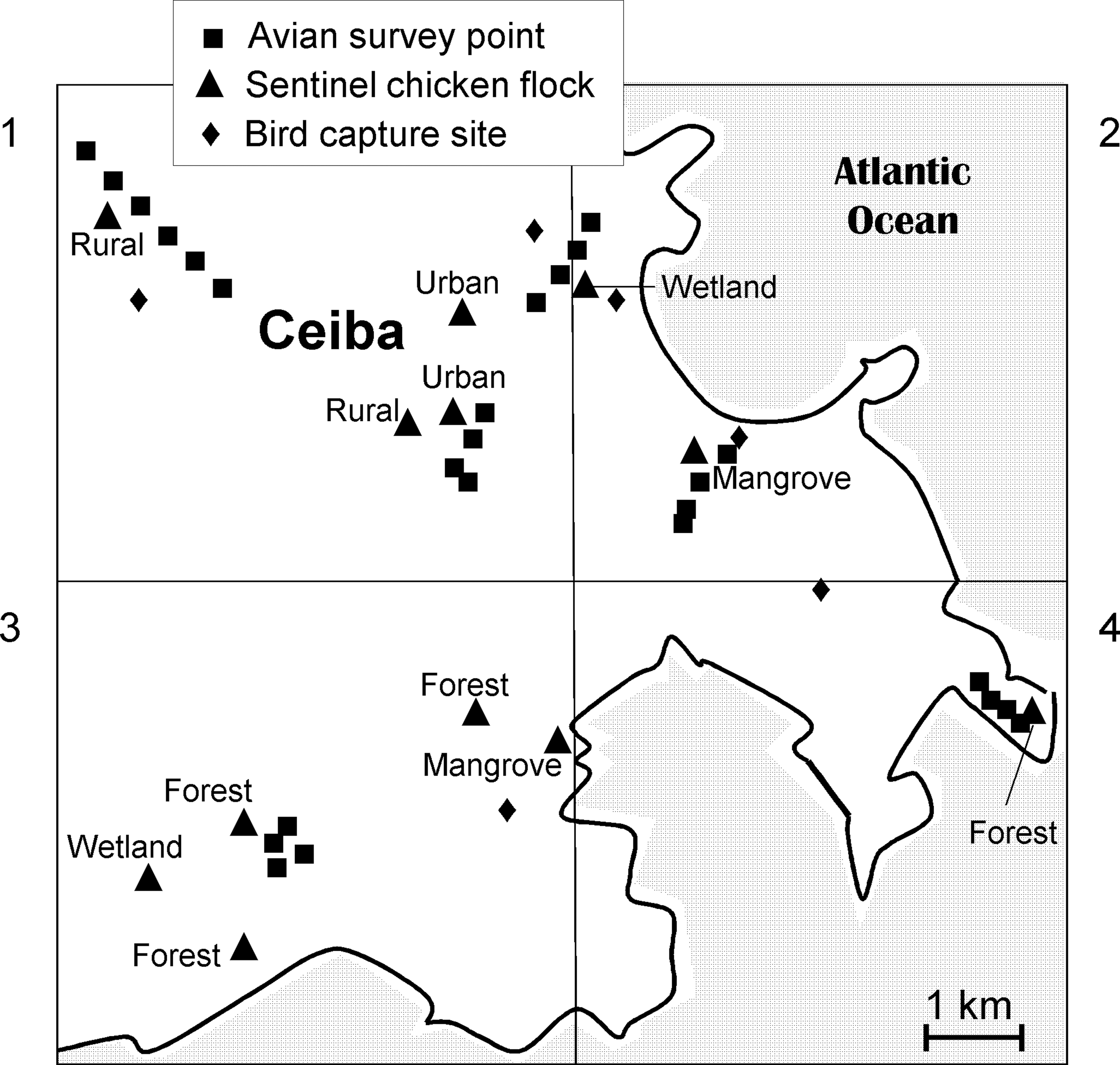
Schematic of the 100-km2 study area, with labels indicating 6 bird capture locations, 26 point count locations for surveying avian relative abundance, and 12 sentinel chicken flocks. The habitat type was classified at each chicken flock location.
Bird sampling
Six locations were selected for free-ranging bird capture based on observations of high bird densities immediately before the bird capture activities. Bird capture and sampling occurred during 20 July—2 Aug 2007, in urban (one location), rural (four locations), and mangrove forest (one location) habitats. Birds were captured by using mist nets (Avinet, Inc., Dryden, NY). Birds were attracted to the mist nets by baiting the ground underneath and in front of the nets with mixed bird seed and moistened dog food pellets. Nets were monitored almost constantly to ensure that birds were not entangled for more than 20 min. Birds were handled with disposable latex gloves. Blood samples were obtained via jugular venipuncture with sterile, disposable 26-g and 27-g needles attached to 1-cc tuberculin syringes. A maximum of 0.65 mL of whole blood or 1% of a bird's mass, determined by using a 100-gram precision spring scale (Avinet, Inc.), was obtained, whichever volume was smaller, and placed into Microtainer® serum separator tubes. Samples were left at ambient temperature for 15–60 min, and then placed on wet ice for the remainder of the day. Each night, samples collected that day were centrifuged for separation of serum, and refrigerated for up to 3 days until transported to the laboratory on dry ice. In the lab, samples were stored at −20°C until thawed for testing. Captured birds were each marked with a uniquely numbered aluminum leg band (provided by the U.S. Geological Survey Bird Banding Laboratory, Patuxent, MD). Data collected for each bird included species, determined with the aid of a local bird identification manual (Raffaele et al. 1998), age (categories included hatch-year, after-hatch-year, second-year, after-second-year, etc.), determined with the aid of a bird banding manual for North American birds (Pyle et al. 1997), sex (categories included male, female, unknown), location, band number, and specimen number for the blood sample. Recaptured birds and birds weighing less than 9 g were released without sampling.
Serologic testing
Serum samples were heat inactivated for 20 min at 56°C. Dilutions of 1:10 and 1:100 were prepared in blocking buffer (5% nonfat milk with 0.1% Tween 20 in phosphate-buffered saline), and screened in triplicate at each dilution for flavivirus group-specific antibodies by using a blocking enzyme-linked immunosorbent assay (ELISA) protocol with the flavivirus group-reactive monoclonal antibody 6B6C-1 (Blitvich et al. 2003). Samples testing positive (using mean 30% inhibition as the criterion for a positive test result) were further evaluated by the same blocking ELISA with the WNV-specific monoclonal antibody 3.1112G. Samples positive in this second test were considered previously infected by WNV. For samples with sufficient quantity, additional testing was performed by using a plaque-reduction neutralization test (PRNT), modified for use at biosafety level 2 (Komar et al. 2009). Criteria for a positive PRNT were 90% neutralization of approximately 100 plaque-forming units of a recombinant yellow fever virus chimera expressing the WNV envelope protein, by serum diluted 1:40, and fourfold greater anti-WNV titer compared with anti-Saint Louis encephalitis virus (SLEV) titer.
Relative abundance estimates
An arbitrary square of side=10 km was selected to define limits of the study area (Fig. 1). Local avian populations were estimated by extrapolation, multiplying each habitat-specific density by the quantity of land assigned to each habitat type (Table 1). Densities were estimated by conducting twenty-six morning point counts of duration=4 min and fixed radius=50 m. Transient flyovers were excluded from the counts, whereas aerial species foraging over the point count circle (i.e., Antillean nighthawk and cave swallow1) were included. Points were selected based on accessibility by road or walking trail, and by uniformity of habitat within each point count circle. Points were separated by at least 150 m. Counts within a habitat type were selected along transects following roads or paths. Habitat types surveyed for avian densities included mangrove forest (4 points), rural (6 points), urban (8 points), evergreen forest (4 points), and semideciduous forest (4 points). Open water and open space were not surveyed, as few birds roost in these habitats at night, when WNV transmission is expected to occur. For the purpose of estimating area-wide population bird populations, evergreen and semideciduous forests were combined. Adelaide's warbler, and perhaps other forest species found primarily in one or the other of the forest types, were, thus, overestimated. Each point was surveyed once, between the hours of 06:15 and 08:55. Surveys were conducted on 3 separate days during the period July 21–27, 2007. Counts were reset at one-min intervals so that each bird had its own “encounter history” for the four 1-min periods of each count. These encounter histories enabled an adjustment of density measurements for species-specific detection biases using closed capture models with Program MARK (White and Burnham 1999). Program MARK also output 95% confidence intervals on abundance estimates within the area surveyed when sample sizes were adequate. The limits of these intervals were adjusted to reflect the extrapolated densities for each habitat and the area-wide population estimates.
Additional statistical analyses
We calculated 95% confidence intervals for seroprevalence proportions by using the Wilson score method (S-PLUS 6.1 Professional software; Insightful, Inc., Seattle, WA). Seroprevalence proportions were compared by using the Fisher exact test or the Pearson χ 2 test. For multiple comparisons, Bonferroni adjustments were applied. The limits of the 95% confidence intervals on the calculated number of infections (product of abundance and seroprevalence) for each bird species was calculated by log transforming the product, applying the method of Zou and Donner (2008), and back transforming the result.
Results
A total of 398 birds were captured and blood sampled. Testing was completed on 366 serum samples from 20 species of birds, with 39 samples from seven species testing positive for WNV-reactive antibodies by blocking ELISA (Table 2). A subset of 30 positive samples, for which sufficient serum was available, was further tested by PRNT. All but one sample (97%) was reactive at titers ≥40; minimum fourfold greater titer compared with corresponding titer for SLEV, a closely related virus that cross-reacts serologically with WNV and circulates in the region, was evident in 25 (83%) samples. Two samples had anti-WNV titers of 40 with anti-SLEV titers of <40, and two had anti-WNV titers ≥320 with anti-SLEV titers either 160 or also ≥320, possibly indicating previous infections with both viruses. The seven species with seropositive individuals were (in descending order by seropositive rate) the red-legged thrush, white-winged dove, pearly-eyed thrasher, northern mockingbird, gray kingbird, Greater Antillean grackle, and house sparrow (Table 2). Seropositivity was evaluated separately for two age classes: “hatch-year” (<1 year old) and “after-hatch-year” (≥1 year old). For each of the seven species with confirmed infections, all exhibited higher infection rates in the hatch-year age class. However, the differences in these rates among species tested individually failed significance tests. Combining all species together, age did have a significant effect (p=0.001). The effect was also observed if the analysis was limited to the seven seropositive species combined (p=0.005).
The category “All ages” includes two individual birds for which age was not determined (northern mockingbird and shiny cowbird).
These include Spotted sandpiper (Actitis macularius, Scolopacidae; sample size=3, upper confidence limit=56.2), common ground-dove (Columbina passerina, Columbidae; 2, 65.8), zenaida dove (Zenaida aurita, Columbidae; 1, 79.4), mangrove cuckoo (Coccyzus minor, Cuculidae; 2, 65.8), Puerto Rican woodpecker (Melanerpes portoricensis, Picidae; 3, 56.2), Caribbean elaenia (Elaenia martinica, Tyrannidae; 8, 32.4), Puerto Rican flycatcher (Myiarchus antillarum, Tyrannidae; 1, 79.4), bananaquit (14, 21.5), Puerto Rican spindalis (Spindalis portoricensis, Thraupidae; 2, 65.8), black-faced grassquit (Tiaris bicolor, Emberizidae; 14, 21.5), shiny cowbird (Molothrus bonariensis, Icteridae; 25, 13.3), nutmeg mannikin (Lonchura punctulata, Estrildidae; 58, 6.2), and pin-tailed whydah (Vidua macroura, Viduidae; 1, 79.4).
Thirty-four bird species were identified during point counts, broken down by habitat, as follows: urban—19 species, forest—18, mangrove—14, and rural—12. Bird density was in the same order of magnitude in all four habitats (roughly 10 individuals per hectare), but urban and mangrove habitats hosted about twice the density of birds than rural and forest habitats (Table 3). Open land (e.g., airport runways, parking lots) and open water were not assessed for bird density. Species richness and passerine bird density were greatest in urban and mangrove habitats. Shannon's diversity index measured roughly the same (range 0.9–1.1) for bird species among all four habitats (Table 3).
Density estimates presented in this table are for multispecies groups and are not adjusted for detection probabilities, and, hence, are conservative estimates; unit of measure is ha−1.
S represents a measure of species richness, the mean number of species per point count.
S-passerine represents the mean number of passerine species per point count.
H represents the calculated Shannon's Diversity Index for all birds.
ha, hectares; SD, standard deviation.
Area-wide population estimates indicated that the Greater Antillean grackle was the most abundant bird in the study area, followed by the black-faced grassquit and bananaquit (Table 4).
Densities are unadjusted for detection probabilities due to insufficient sample sizes for most species.
Former family assignment. Current status is Genus Incertae Sedis.
NA, not available.
To infer the relative number of WNV infections per avian species within the study area, we multiplied the relative abundance (in units of birds per study area) by the prevalence of WNV-reactive antibodies (no units) to derive an estimate of the number of WNV-infected birds within the study area, for each of the seven species found to have been exposed to some degree (Table 5).
The inferred number of West Nile Virus-infected birds in the study area is calculated as the product of population and the infection rate.
Area-wide population estimates and their confidence intervals are extrapolated from habitat specific densities, adjusted for detection probabilities, which were determined by Program MARK and the estimated areas of each habitat presented in Table 4.
Since mortality has not been observed for these species in Puerto Rico, seroprevalence is a surrogate for infection rate. Values come from the percentages in Table 2 for all age classes combined.
Population of red-legged thrush was set at 92, one below the minimum detectable number from point count surveys in the rural habitat.
95% CI, 95% confidence interval.
Discussion
A focus of intense WNV transmission provided an opportunity to investigate the avian hosts of WNV in Puerto Rico. Previous information on avian WNV hosts in the Caribbean region derived from expeditions to search for WNV-infected birds in the absence of documented local transmission (DuPuis et al. 2003, 2005, Komar et al. 2003b, 2005b). WNV first appeared in this region in a human resident of the Cayman Islands in 2001 (Komar and Clark 2006). In early 2002, a survey of 348 Jamaican birds detected specific WNV-neutralizing antibodies in 4 white-chinned thrush (Turdus aurantius, Turdidae), 2 each of Jamaican elaenia (Myiopagis cotta, Tyrannidae), bananaquit, and black-faced grassquit, and 1 each of 7 other species (DuPuis et al. 2003). More seropositive birds were discovered among 189 resident birds in the Dominican Republic in 2002 and 2003, including 4 black-crowned palm-tanager (Phaenicophilus palmarum, Thraupidae), 2 each of village weaver (Ploceus cucullatus, Ploceidae), Hispaniolan lizard-cuckoo (Saurothera longirostris, Cuculidae), Greater Antillean bullfinch (Loxigilla violacea, Emberizidae) and red-legged thrush, and 1 each of 5 other species (Komar et al. 2003b, 2005b). In 2004, 2 more seropositive red-legged thrush (of 364 resident birds sampled) were detected in Cuba, and 1 bananaquit (of 710 resident birds sampled) in Puerto Rico was seropositive (DuPuis et al. 2005). Detection of WNV-reactive antibodies in these studies signaled past infections. Most of these seropositive birds were classified as after hatch-year; and thus, there was little evidence of current transmission. Whether the transmission events that led to these infections were focused in time or cumulative over a long period remains unknown.
In our study, we targeted a region with temporally focused WNV transmission. Further, we estimated relative abundance of birds. Although uncommon birds may be infected and may transmit viruses, only abundant birds will be important amplifying hosts (Scott 1988). Passerine birds tend to be highly competent for infecting mosquitoes with WNV (Komar et al. 2003a), and the five most abundant birds in the study area were all passerine: Greater Antillean grackle, black-faced grassquit, bananaquit, gray kingbird, and house sparrow. WNV-reactive antibodies indicated that at least three of these were frequently infected (grackle, kingbird and sparrow) but not the other two (sample sizes, n=14 each), although seropositive black-faced grassquit and bananaquit have been previously detected. However, our calculations of the approximate number of infected birds in our study area also indicated substantial transmission of WNV to the white-winged dove, pearly-eyed thrasher, northern mockingbird, and, to a lesser extent, red-legged thrush. Of these, only the white-winged dove is non-passerine, and, consequently, is likely to be less competent to infect blood-feeding mosquitoes. However, other studies (e.g., Kent et al. 2009) have demonstrated that doves may contribute significantly to the mass of WNV-infectious mosquitoes in some regions; and therefore, this species should not be ruled out as potentially important as an amplifying host.
We compared infection rates in hatch-year birds to those of older birds of seven species to test the hypothesis that seroprevalence rates in the older birds were elevated compared with those in the hatch-year birds, a pattern that would be expected if WNV was enzootic. None of these seven comparisons supported this hypothesis; and therefore, local WNV activity before 2007 was presumed negligible. Conversely, hatch-year birds were more frequently infected, as observed elsewhere (Hamer et al. 2008). This finding is difficult to interpret beyond disproving our hypothesis. Factors contributing to the greater seroprevalence among hatch-year birds may include, among others, vector feeding preference on hatch-year birds, antibody decay in older birds infected during previous years, age-dependent survival of WNV infection, and age-dependent anti-mosquito defensive behavior.
The Greater Antillean grackle stood out from the other species as being the most frequently infected bird species in the study area. Although its competence for transmitting WNV to feeding mosquitoes remains untested, other grackle species are highly competent as vertebrate reservoir hosts for WNV (Komar et al. 2003a, Guerrero-Sanchez et al. 2011). The Greater Antillean grackle utilizes many habitats, often near human settlements, throughout Puerto Rico and elsewhere in the Greater Antilles. The grackles nest and roost communally, and presumably, thus, attract large numbers of host-seeking Culex quinquefasciatus. Communal roosts of passerine birds have been implicated as WNV amplification foci in the United States (Kent et al. 2009, Diuk-Wasser et al. 2010). Bloodmeal identification studies have reported Culex bloodmeals derived from grackles in Colorado and Texas (Molaei et al. 2007, Kent et al. 2009).
The house sparrow is primarily an urban species, so this species is among the leaders of infected birds in the urban zone. Behavior may enhance this species' involvement, as these birds also nest and roost communally. In experimental infection studies, house sparrows are strong amplifiers of WNV, regardless of the strain of virus evaluated. The house sparrow's great amplifying potential is not predicted by its seroprevalence, which was 6% among hatch-year birds, lowest among the seven seropositive species. The house sparrow is the only one of these seven species known to die from infection with the New York 1999 strain of WNV (Komar et al. 2003a). The susceptibility of sparrows to fatal infection from the Puerto Rican strain remains unknown, but mortality in this species would contribute to a low detection rate for seropositive individuals. To calculate the number of infected sparrows assuming a mortality rate, m, of 50% (a value experimentally derived; Komar et al. 2003a), we calculate infection rate, I, based on mortality and seroprevalence, s, from the following equation (Komar et al. 2005a):
Using s=0.05 and m=0.5, we find that I=0.095, and the inferred number of infected sparrows increases from 217 to 414. Therefore, if WNV-attributed mortality in sparrows is passing unnoticed, then our calculations underestimate the potential importance of these birds as reservoirs in urban habitat.
The pearly-eyed thrasher and red-legged thrush are two additional passerine species that may be amplifying hosts. Although the thrush was not commonly observed (or captured) in our study site, its infection rate was very high (80%). The red-legged thrush is very common in gardens of suburban San Juan, and, thus, should be considered a candidate important amplifying host in Puerto Rico and other Caribbean locations where it occurs (i.e., Hispaniola, Cuba, Cayman Islands). WNV antibody-positive red-legged thrushes have been detected in Cuba and the Dominican Republic (Komar et al. 2003b, DuPuis et al. 2005). Other congeners (e.g., American robin, Turdus migratorius; clay-colored thrush, T. grayi, etc.) have been reported WNV antibody-positive in other geographic regions such as Jamaica, United States, Guatemala, Venezuela and Argentina (DuPuis et al. 2003, Kilpatrick et al. 2006, Bosch et al. 2007, Diaz et al. 2008, Morales-Betoulle et al. 2008).
Our study adds to a growing literature on the ecology of WNV in Caribbean ecosystems, identifying candidate WNV-amplifying hosts, particularly Greater Antillean grackle, gray kingbird, and, in urban areas, house sparrow. Competence studies utilizing the local WNV strain in birds representing local populations and bloodmeal identification studies are needed to pinpoint the vector/host interactions responsible for transmission. Nocturnal communal roosts of grackles and sparrows should be evaluated as potential transmission hot spots, where anti-vector interventions could be focused in the event of an epidemic threat to humans or horses. Avian mortality due to the Puerto Rican WNV strain should be assessed to determine whether bird carcass surveillance for WNV would be useful, and whether the numerous threatened populations of birds on Caribbean islands are at risk. Finally, the relationship between amplification of WNV by birds (and/or other vertebrate hosts) and risk of human illness in Puerto Rico and elsewhere in the Caribbean Basin should be determined.
Footnotes
Acknowledgments
Numerous private property owners granted permission for field studies, in particular the managers of the Roosevelt Roads Naval Base. Veronica Acevedo, Heidi Acosta, Manuela Beltran, Kali Shaw, and others provided field assistance and laboratory support. Josh Dooley assisted with data analysis. Mark Delorey provided statistical advice. Roberto Barrera provided the aerial image of the study region. Animals were handled in this study following the guidelines of the Public Health Service and National Research Council, and approved by the Institutional Animal Care and Use Committee of CDC-DVBD. This work was funded by the Centers for Disease Control and Prevention. Support for employees of the Puerto Rico Department of Health was administered through a Cooperative Agreement. The statements and opinions expressed in this article are those of the authors and do not necessarily represent the official policy of the Centers for Disease Control and Prevention or the U.S. Government or the Government of the Commonwealth of Puerto Rico.
Disclosure Statement
No competing financial interests exist for any of the authors.
1
Scientific names for these bird species are provided in the data tables for these and all other species mentioned in the remainder of the text. Scientific names follow current nomenclature of the American Ornithological Union.