
Abstract
Culex torrentium and Culex pipiens are sibling species and potential viral vectors that coexist in Europe. Larvae and females of the two species are morphologically almost identical, and reliable identification can only be done on males. To investigate the distribution and relative abundance of the two species in Sweden, we collected Culex larvae from sites spread over the country, identified them as Culex pipiens/torrentium based on morphology, and identified them to species using a recently developed restriction enzyme method. Cx. torrentium was the dominant species (89%, n=1012) and it occurred in 48 of the 49 sites investigated, and also dominated in most of the study sites. The proportion of Cx. pipiens larvae in relation to Cx. torrentium collected at each site decreased with both increasing latitude and altitude, and the presence of Cx. pipiens decreased with latitude. In addition, Cx. pipiens/torrentium females were sampled with Centres for Disease Control light traps baited with carbon dioxide. The overall country mean was 4.0 Cx. pipiens/torrentium caught per trap night, with decreasing numbers of Cx. pipiens/torrentium caught per trap night with increasing latitude. Thus, the abundance of Cx. pipiens/torrentium decreased, but the proportion Cx. torrentium increased, with increasing latitude. This is the first study that shows the vast dominance of Cx. torrentium over Cx. pipiens in Sweden. The unexpected dominance of Cx. torrentium highlights the importance of distinguishing between the two species in studies of Culex-borne arboviruses in Europe.
Introduction
Cx. pipiens is the most well known of the two species, and it is one of the suggested vectors of West Nile virus (Flaviviridae; Flavivirus) in Europe (Savage et al. 1999, Hubálek et al. 2000, Esteves et al. 2005, Fyodorova et al. 2006). In transmission experiments, it has shown moderate vector competence for West Nile virus (Balenghien et al. 2008), and for Sindbis virus (Togaviridae: Alphavirus) (Lundström et al. 1990a, 1990b). Sindbis virus is a mosquito-borne and bird-associated zoonosis that causes outbreaks of rash and arthralgia in humans in Scandinavia and Russia (Lundström 1999). In Sweden, Sindbis virus has been isolated with the highest field infection rate from unidentified Cx. pipiens/torrentium (Francy et al. 1989). It has also been isolated more recently in Germany from a pool including both Cx. torrentium and Cx. pipiens (Jöst et al. 2010).
Cx. torrentium is much less well known than Cx. pipiens and has only been tested for its vector competence for Sindbis virus. It proved to be an extremely efficient vector that was very susceptible to oral infection and a highly competent transmitter (Lundström et al. 1990a, 1990b). Based on its high vector competence, the potential virus isolates and that it mainly seems to feed on birds (Service 1968b, 1971), Cx. torrentium is considered to be the main enzootic vector of Sindbis virus in Sweden.
Both Cx. torrentium and Cx. pipiens occur in Sweden, but just like in the rest of Europe, their distribution and abundance is largely unknown since only scattered information based on identified males is available (Jaenson and Niklasson 1986, Andersson and Jaenson 1987, Francy et al. 1989, Lundström et al. 1990a). Recently, we developed a molecular method, based on reliably identified males of each species, to identify the two species accurately independently of life stage and sex (Hesson et al. 2010). We use this method to investigate the geographic distribution and relative abundance of Cx. torrentium and Cx. pipiens in Sweden.
Materials and Methods
Larval collections
Larvae of Cx. pipiens/torrentium were collected during four summers (2006:1 site, 2007:1 site, 2008:18 sites, 2009:29 sites) mainly in mid-July to mid-August (and at two occasions in September), at 49 sampling sites (Fig. 1). The sampling sites were distributed throughout Sweden, excluding the Scandinavian mountain range in the northwestern parts of the country since previous sampling efforts have shown that Culex mosquitoes are difficult to find in these areas (Schäfer and Lundström 2001). The larval sampling sites were both artificial (e.g., car tires and water barrels) and natural (e.g., ditches and puddles). Each sampling site was represented by one pool of water, and the population of Cx. pipiens/torrentium larvae present there. Collected larvae were killed and preserved in 70% ethanol in the field, and were later morphologically identified to Cx. pipiens/torrentium using a stereo microscope. Identification of larvae was based on Becker et al. (2003).
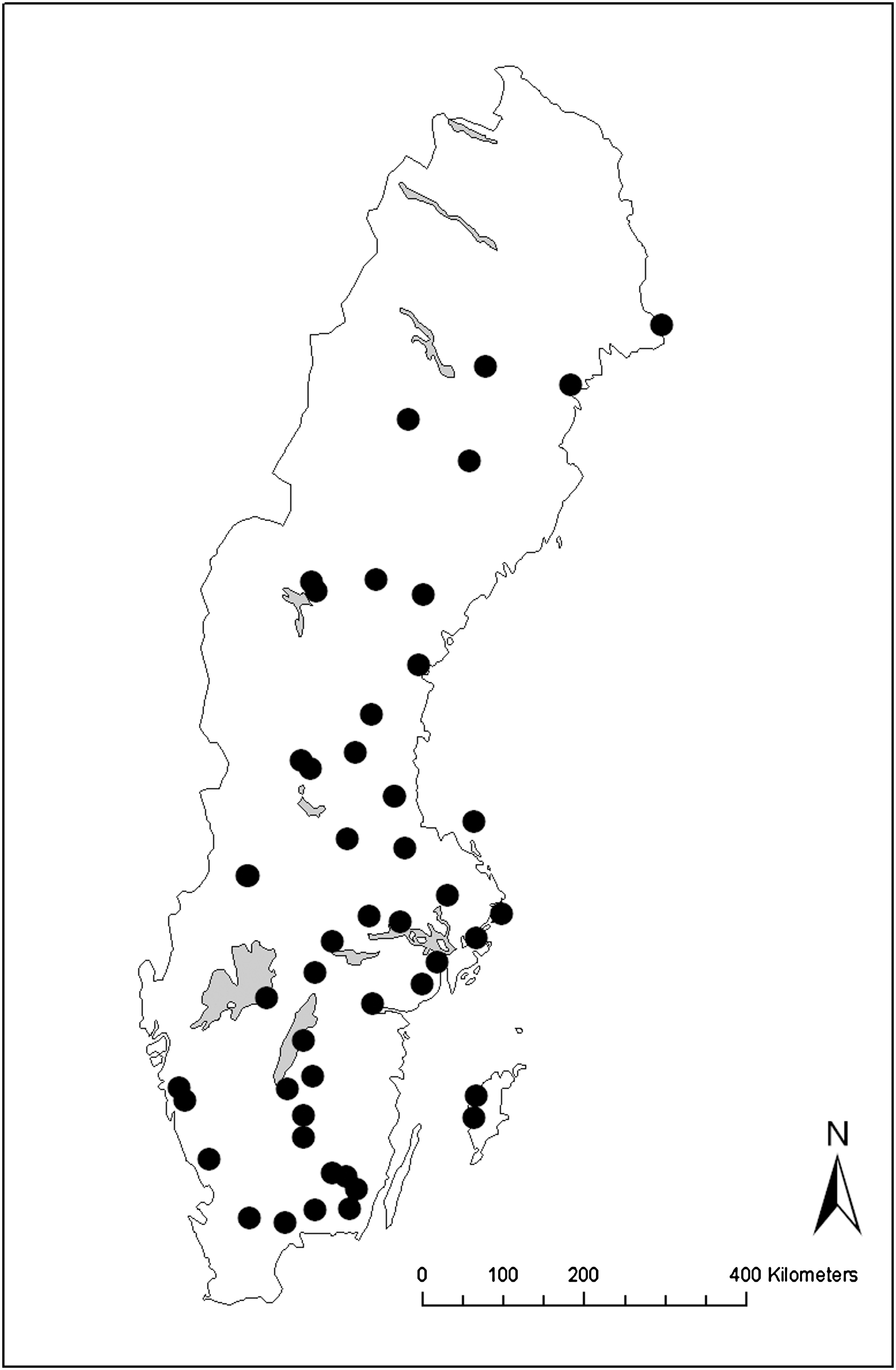
Distribution of the 49 sampling sites for Culex pipiens/torrentium larvae in Sweden. The majority of sampling sites were visited in 2009 (59%) and 2008 (37%), whereas a few sites were visited in 2006 (2%) and 2007 (2%).
Adult collections
Data and samples of adult Cx. pipiens/torrentium were extracted from the Swedish mosquito-monitoring program at 72 sites distributed in south, central, and north Sweden (Lundström, unpublished). This sampling was performed for five summers between 2005 and 2009. There was local variation in the sampling effort, with the biggest sampling effort concentrated toward central east Sweden. The adult mosquitoes were sampled using Centres for Disease Control miniature light traps (CDC-traps) baited with carbon dioxide. The traps were put out at about 1.5 m above ground level in late afternoon and retrieved the following morning. The mosquitoes were killed by freezing on dry ice (solid carbon dioxide), and stored at −70°C until morphological identification to Cx. pipiens/torrentium using a stereo microscope. Identification of females was based on Becker et al. (2003).
Molecular identification to species
Molecular identification was performed according to the method developed by Hesson et al. (2010). DNA from individual specimens was extracted and part of the mitochondrial CO1-gene was amplified by polymerase chain reaction (PCR), using the primers C1-J-2183 and TL2-N-3014. The PCR products were digested with the restriction enzymes FspBI and SspI (Fermentas), and observed on an agarose gel. The enzyme FspBI cut PCR products of Cx. torrentium into two fragments, whereas PCR products of Cx. pipiens remain uncut. The enzyme SspI cut PCR products of Cx. pipiens into two fragments, whereas PCR products of Cx. torrentium remain uncut.
The molecular identification was performed on 16–25 larvae from each sampling site, with the exception of Piteå-07, where 45 larvae were analyzed. In addition, 199 adult females were also molecularly identified to species. These females were selected from mosquito collections from 46 sites within the Swedish monitoring program (Lundström, unpublished). However, only samples from south and central Sweden were available for identification.
Statistical analyses of the geographical distribution and relative abundance of Cx. torrentium and Cx. pipiens
Longitude and latitude were recorded, for each larval and adult sampling site, using a hand-held GPS (Magellan eXplorist 600; Magellan). Additional information on altitude, mean temperature, and cumulative precipitation during May to August was extracted from WorldClim, which is based on data from 1950 to 2000 (
First, we tested for differences in abundance of the two species in all larval samples with a paired t-test. Then, we calculated the proportion (relative abundance) of each species (i.e., between 0 and 1) at each sampling site. To study how the proportional abundances of the two species were associated to the explanatory variables (longitude, latitude, altitude, cumulative precipitation, mean temperature, year, and day of sampling), we used Proc Genmod in SAS 9.1.3 (SAS Institute 2003) with a logit link function and binomial distribution. Here, we use the proportion of Cx. pipiens as our dependent variable. Explanatory variables that had p-values higher than 0.1 were removed in a backward manner until all explanatory variables in the model had p<0.1.
For Cx. pipiens larvae we then investigated how the presence or absence changed with the explanatory variables (if Cx. pipiens was present in a sample it was scored as “1,” if not as “0”) by using a similar generalized mixed model as above. Explanatory variables were removed in a backward manner until all remaining variables in the final model had p<0.1. This analysis was not conducted for Cx. torrentium because it occurred in all samples but one (see Results section).
For adult samples, we tested how the presence or absence of each species changed with the explanatory variables. The analysis was based on a generalized mixed model using the GLIMMIX macro (Wolfinger and O'Conell 1993) in SAS 9.1.3 (SAS Institute 2003), with a logit link function and binomial distribution. The repeated sampling over years at some sites was modeled with sampling site as repeated factor. Explanatory variables were removed in a backward manner until all variables had p<0.1.
Statistical analysis of the total abundance of Cx. pipiens/torrentium
From the 5 years of adult mosquito monitoring we extracted the number of Cx. pipiens/torrentium (not identified to species) caught per CDC-trap and night. To study how the total abundance of these Culex species varied over Sweden, we used a mixed model (PROC MIXED) in SAS 9.1.3 (SAS Institute 2003). Abundance was measured on a weekly basis each year, and was estimated as the total number of Culex in a trap divided by number of trap nights. We used the natural logarithm of the number of Cx. pipiens/torrentium trapped per night as a dependent variable. As explanatory variables we used year as a fixed category variable, week and week2 as fixed continuous variables, and latitude and longitude as random variables. To account for the repeated sampling over years, the sampling locality was entered as a repeated factor.
Results
Geographical distribution and relative abundance of Cx. torrentium and Cx. pipiens
In total, 1012 larvae from 49 sampling sites were identified to species. Overall, Cx. torrentium dominated samples (89%) over Cx. pipiens (11%) (t 1,47=13, p<0.001), and occurred in all but one site. At 19 sites both species occurred together. Examples of these habitats were garden water barrel, garden pond, car tire, manure water, concrete tube, tractor scoop, water puddle in forest, and bathtub for animal drinking. Cx. torrentium was observed alone in 29 of the 49 sites, and none of these sites appeared unique as compared to sites with both species present. Also, Cx. torrentium was the dominant species in all samples except three, where Cx. pipiens constituted 100%, 65%, and 55%, respectively. The proportion of Cx. pipiens larvae (in relation to Cx. torrentium) decreased with increasing latitude (χ 2 (1)=13, p<0.001, n=49) and with increasing altitude (χ 2 (1)=8.2, p=0.004, n=49). No Cx. pipiens larvae were found above 227 m, although sampling was done up to 452 m.
The presence of Cx. pipiens larvae decreased with increasing latitude (χ 2 (1)=3.9, p=0.047, n=49). Since Cx. torrentium was found at almost all sites, we considered a similar analysis for Cx. torrentium to be noninformative.
In addition, we species-identified 199 adults from 46 sites in our monitoring program situated in south and central Sweden (none above 110 m altitude). We identified 42 (21%) of them as Cx. torrentium and 157 (79%) of them as Cx. pipiens. The presence of adult Cx. pipiens decreased with increasing latitude (F 1,41=5.7, p=0.02), whereas the presence of adult Cx. torrentium increased with latitude (F 1,41=7.5, p=0.009). A summary of all species identified larvae and adults is shown in Table 1. To make comparisons possible, we display the data by vegetation zones, since our study sites include four of the six vegetation zones in Sweden.
Absolute numbers in parentheses, and absence of sampling is indicated by n/a. (For a descriptive map of the vegetation zones, see
In both the larval and the adult material, we found that latitude and/or altitude explained most of the variation in the presence of Cx. pipiens. Notably, there was some covariation between latitude/altitude and other explanatory variables, such as temperature. Temperature was negatively correlated with both latitude (r=−0.22) and altitude (r=−0.52). Thus, the geographic distribution of Cx. pipiens was associated to temperature. We mapped all larvae and adults identified as Cx. torrentium (Fig. 2) and Cx. pipiens (Fig. 3). These figures indicate the temperature range where Cx. torrentium was found but not Cx. pipiens. We marked the lowest mean temperature in May to August for a site where Cx. torrentium was present (10.5°C), and the corresponding temperature for a site where Cx. pipiens was present (11.9°C). We also found that the sampling sites with the lowest minimum temperatures in May to September were consistently sites where Cx. torrentium was present (Table 2).
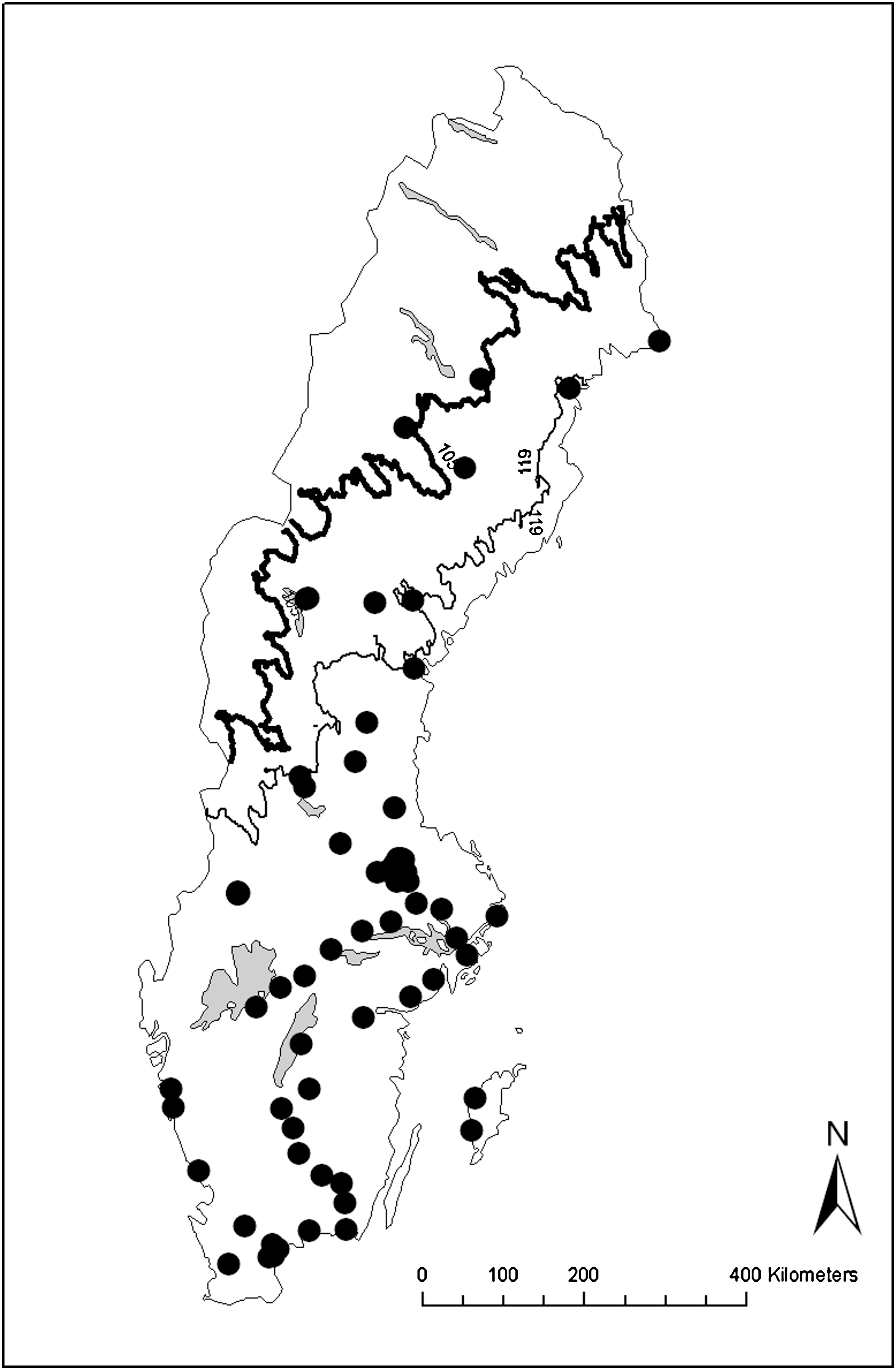
Distribution map for Cx. torrentium in Sweden, based on molecularly identified larvae and adults. Marked with a thick line is the lowest mean temperature (May–August 1950–2000, WorldClim) for a site where Cx. torrentium was found (10.5°C). The thinner line represents the corresponding temperature for a site where Cx. pipiens was found (11.9°C).
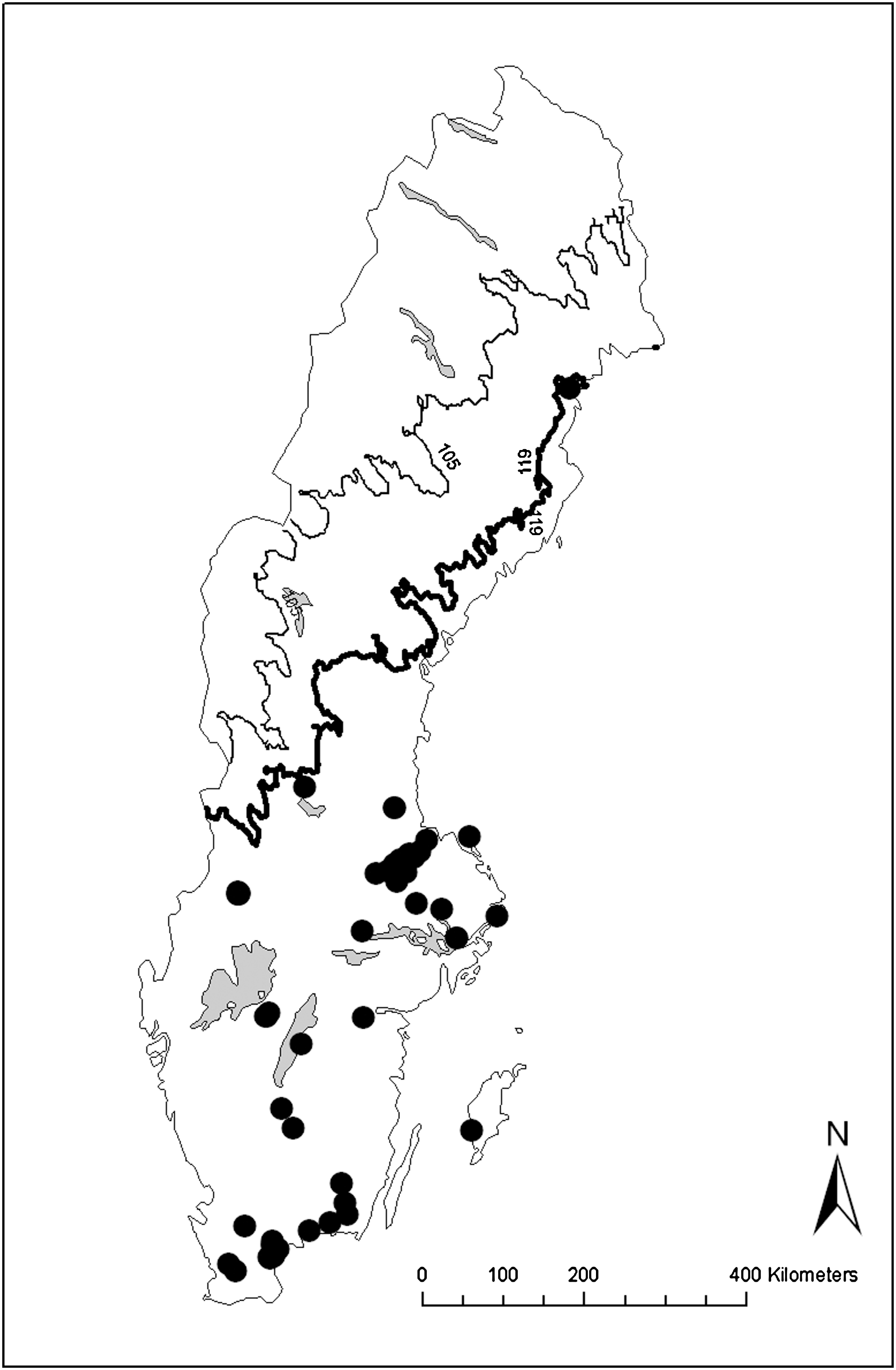
Distribution map for Cx. pipiens in Sweden, based on molecularly identified larvae and adults. Marked with a thick line is the lowest mean temperature (May–August 1950–2000, WorldClim) for a site where Cx. pipiens was found (11.9°C). The thinner line represents the corresponding temperature for a site where Cx. torrentium was found (10.5°C).
Total abundance of Cx. pipiens/torrentium
In total, 6287 Cx. pipiens/torrentium females were sampled over 1546 trap nights. The overall country mean was 4.0 Cx. pipiens/torrentium caught per trap and night, with variations between regions (south: 44, central: 0.86, north: 0.24). Thus, the number of Cx. pipiens/torrentium caught per trap night decreased with increasing latitude (F 1,64=547, p≤0.001, partial r 2=0.25). This effect was found for all years investigated except for 2005, where there were no observations on different latitudes.
In 2007, extraordinary large amounts of Cx. pipiens/torrentium females were caught in the south of the country, with up to 1434 Cx. pipiens/torrentium per trap and night at one site. Species identification of 60 individuals from catches in this area in 2007 showed that the majority (92%) were Cx. pipiens.
Discussion
In our larval collections, Cx. torrentium was more common than Cx. pipiens in Sweden. It occurred from the very south to the very north of the country and at all sampled altitudes (0–452 m). In contrast, published literature on Cx. torrentium is very scarce and much of the information available is anecdotal. Initially, it was believed to be a clean water species, but this view was revised when the larvae were found in a variety of aquatic habitats, including ponds rich in organic content and sometimes together with Cx. pipiens (Scherpner 1960, Gillies and Gubbins 1982, Ishii and Sohn 1987, von Struppe 1989, Raymond 1995). Based on a few empirical studies, Cx. torrentium is known as a species adapted to cold habitats (von Struppe 1989), and to occur at high altitude with males caught at 1500 m in the Pyrénees (Sicart 1954). However, a more recent study found that Cx. torrentium larvae occurred in Germany at a mean water temperature of 24°C, ranging between 21.3°C and 33.7°C (Küpper et al. 2006). These rather high temperatures show that Cx. torrentium, besides being cold tolerant, also can persist under warmer conditions. The upper limit of Cx. torrentium's temperature range, and its southern geographical limits, are not known, but the species has been found in Southern Europe, Iran, and Iraq (Harbach 1988). Within Sweden, we see no southward decrease in the presence of Cx. torrentium.
Adult identification was performed mainly on individuals caught in the two most southern vegetation zones in Sweden (Table 1), and resulted in more Cx. pipiens identified than Cx. torrentium. These numbers might to some extent reflect the skewed species identification effort of adult mosquitoes to south and central Sweden, where both Culex mosquitoes in general and Cx. pipiens are more abundant. However, it might also be an effect of sampling bias. The larval habitats we sampled were all well documented to be appropriate for both species. Adult attraction to CDC-traps baited with carbon dioxide has, however, been demonstrated to differ also in previous studies. In Germany, catches from ovitraps and carbon dioxide-baited traps in the same area were compared, based on molecular identification of species. The ovitraps caught 55% Cx. pipiens and 45% Cx. torrentium, whereas the carbon dioxide-baited traps caught 96% Cx. pipiens and only 4% Cx. torrentium (Weitzel et al. 2011). A study in Luxemburg, using the same molecular method of identification, found that carbon dioxide-baited traps caught 18 Cx. pipiens, and 1 Cx. torrentium, whereas larval sampling found 2 Cx. pipiens and 11 Cx. torrentium (Beck et al. 2003). This is in line with several studies pointing out that CDC light traps baited with carbon dioxide do not correctly represent abundance for many mosquito species (DiMenna et al. 2006, Giberson et al. 2007), and also highlights the question if these traps really give a fair picture of the relative abundance between Cx. pipiens and Cx. torrentium.
Both the presence of Cx. pipiens larvae, and its proportions in relation to Cx. torrentium, decreased with altitude and no Cx. pipiens was found above 227 m. For adults, all samples were collected below 200 m, and no significant effect of altitude could be detected. However, it was significant that the presence of Cx. pipiens adults decreased with latitude. Previously, Cx. pipiens males have been found at 1500 m in the Pyrenées (Sicart 1954), which is the same as for Cx. torrentium. Thus, high altitude per se is not a limiting factor. Rather, Cx. pipiens' limited distribution in Sweden is likely to be caused by underlying factors connecting altitude and latitude, such as temperature. We found that air temperature was negatively correlated with both altitude and latitude and thus, as expected, positively correlated with the presence of Cx. pipiens. In addition, the sites were Cx. pipiens larvae were found had higher minimum temperatures than sites where Cx. torrentium was found. We cannot specify what factors limit Cx. pipiens at higher altitudes or latitudes, but some temperature constraints seem likely. This needs to be further investigated since, within Sweden, it may exist at the lower end of its temperature range.
In Europe, the research community has been focused on Cx. pipiens, and it has been suggested that this species is a vector of West Nile virus, based on virus isolations and experimentally proven moderate transmission capacity (Esteves et al. 2005, Balenghien et al. 2008). In Sweden it was most abundant in the south, where also the highest number of Culex females per trap night was recorded. Cx. torrentium has generally been overlooked both concerning its vector status and abundance. It proved a much more efficient vector than Cx. pipiens for Sindbis virus (Lundström et al. 1990a, 1990b), and whether it is also an enzootic vector for West Nile virus remains to be studied, as transmission experiments are lacking but much needed. The present study makes it reasonable to believe that the former isolates of Sindbis virus, made from a mixture of unidentified Cx. pipiens/torrentium collected in central Sweden (Francy et al. 1989), originated from Cx. torrentium.
In countries where thorough investigations of Cx. torrentium have been made (England, Germany, and Sweden), it has been found to be just as common, or even more common than Cx. pipiens (Jupp 1979, Gillies and Gubbins 1982, von Struppe 1989, Küpper et al. 2006, Hutchinson et al. 2007, Weitzel et al. 2011). This indicates that Cx. torrentium might be more common also in other countries where such studies have not yet taken place. This could be because there is an increased risk of overestimating Cx. pipiens by morphological identification, since the females are identified by a character (prealar scales) missing from Cx. torrentium (rather than the presence of a unique character). The prealar scales often fall off and are sometimes even missing from fresh specimens; thus, Cx. torrentium is easily mistaken for Cx. pipiens (Service 1968a).
We found that Cx. torrentium and Cx. pipiens to a large extent have overlapping geographic distributions in Sweden, and exist in the same habitats. Sporadic reports indicate that Cx. torrentium has a wide geographic distribution in Europe and we suggest it is time to correct the assumptions that the species occurs mainly in cold habitats and on high altitudes. Accordingly, the distribution and abundance of these vector species should be thoroughly revised based on reliably identified field material.
Footnotes
Acknowledgments
We are very grateful to all the people who have helped us with collecting mosquitoes in the field: Anna Hagelin, Uwe Dauernheim, Thomas Persson Vinnersten, Pernilla Wahlqvist, Arne Halling, Hanna Novakovic, Inger Carlsson, Karin Kjellberg, and Clas Ahlm. We also would like to thank Helge Ax:son Johnsons Stiftelse for financial support.
Disclosure Statement
No competing financial interests exist.