
Abstract
Usutu virus (USUV) was discovered in South Africa in 1959. Since then, it has been reported in several African countries including Senegal, Central African Republic, Nigeria, Uganda, Burkina Faso, Cote d'Ivoire, and Morocco. In 2001, USUV has been identified for the first time outside of Africa, namely in Europe, where it caused a significant mortality among blackbirds in Vienna, Austria. In 2009, the first two human cases of USUV infection in Europe have been reported in Italy, causing encephalitis in immunocompromised patients. The host range in Africa includes mainly Culex mosquitoes, birds, and also humans with one benign and one severe case. Given its role as a potential human pathogen and the similar appearance compared with other emerging arboviruses, it is essential to investigate the natural history and ecology of USUV in Africa. In this regard, we review the emergence of USUV in Africa, summarizing data about isolations, host range, and potential vectors, which should help to improve our understanding of the factors underlying the circulation of USUV in Europe and Africa.
Introduction
Another example of an emerging arboviral disease is Chikungunya virus (CHIKV), which has caused large outbreaks in the Indian Ocean Islands and India since 2004 (Kariuki Njenga et al. 2008). Up to 2010, more outbreaks in Indonesia, La Reunion Island, China, and Madagascar demonstrate its continuous activity and emphasize its role as an emerging infectious disease (ProMEDmail 2010). CHIKV originates in Africa and spread to Asia and India (Sudeep and Parashar 2008). In La Reunion Island, CHIKV acquired a selective advantage for the transmission by an alternate mosquito vector. Subsequently, the CHIKV outbreak in Italy in the year 2007 indicates its potential to spread and establish in Europe (Schuffenecker et al. 2006).
Similarly, Usutu virus (USUV), a virus reported to be circulating only in Africa, emerged in 2001 in Europe, causing a significant number of deaths in blackbirds (Turdus merula) (Weissenbock et al. 2002). Even though avian infections were not reported to be fatal in Africa, USUV has been shown to be highly pathogenic to several bird species in Austria, such as the blackbird (T. merula), the great gray owl (Strix nebulosa), the blue tit (Parus coeruleus), the house sparrow (Passer domesticus) (Weissenbock et al. 2003), the great tit (Parus major), the nuthatch (Sitta europaea), the robin (Erithacus rubecula), and the song thrush (Turdus philomeus) (Chvala et al. 2007). The observed symptoms included hepatosplenomegaly, neuronal necrosis, and necrotic changes of liver, spleen, and heart (Chvala et al. 2004). As for WNV in northern America, USUV overwintered in Austria by adapting to local mosquito species and established a transmission cycle among local birds and mosquitoes mainly of the Culex species (Weissenbock et al. 2003, Brugger and Rubel 2009). Despite continuing virus circulation after its occurrence in 2001, a decrease in bird mortality has been recognized, probably resulting from the establishment of herd immunity in bird hosts. Wild bird surveillance indicates seroconversion in a broad range of wild birds in Austria (Meister et al. 2008), suggesting that USUV is already endemic in mosquito and bird species. In addition, USUV has been identified as a cause of death of blackbirds (T. merula) in Hungary (Bakonyi et al. 2007), Switzerland (Steinmetz et al. 2011), and Italy (Busani et al. 2010), in addition to the detection of the virus in great gray owls (S. nebulosa) and boreal owls (Aegolius funeris) in northern Italy (Manarolla et al. 2009). Further, USUV was isolated from Culex pipiens mosquitoes in Spain (Busquets et al. 2008) and Italy as well as from Aedes albopictus mosquitoes in Italy (Tamba et al. 2010). Serological evidence of its circulation has been reported among blackbird, carrion crow (Corvus corone), magpie (Pica pica), and turkey (Meleagris gallopavo) (Buckley et al. 2003) in the United Kingdom, sentinel chicken in the United Kingdom (Buckley et al. 2006) and Italy (Lelli et al. 2008), white stork (Ciconia ciconia), common redstart (Phoenicurus phoenicurus), and osprey (Pandion haliaetus) in Germany (Linke et al. 2007), eurasian coot (Fulica atra) in the Czech Republic (Hubalek et al. 2008), and black-headed gull (Larus ridibundus) in Poland (Hubalek et al. 2008). Moreover, USUV antibodies were detected in horses in Italy (Lelli et al. 2008).
In September 2009, the first two human USUV infections in Europe have been reported in Italy, representing the first cases of human neuroinvasive infection ever, even though neuroinvasiveness in mice after experimental infection has been already demonstrated (Weissenbock et al. 2004). The patients, an orthotropic liver transplantation recipient and a patient with diffuse large B-cell lymphoma, suffered from severe neurological impairment caused by encephalitis, demonstrating the role of USUV as a human pathogen at least for immunocompromised individuals (Cavrini et al. 2009, Pecorari et al. 2009).
Because of the mortality among some bird species after its introduction into Europe, further attention was given to USUV, leading to the availability of more information. However, in Africa, although the virus was identified in the late 1950s, only very limited data have been published. Given the potential of further geographic expansion, changes in the host range (including humans), and the possible modification of biological features of USUV in Europe or other regions of the world, we review in this article the ecology of USUV in Africa to understand more completely the patterns and processes underlying its emergence and identify the gaps in our knowledge.
Discovery and Description of the Virus
USUV was recovered for the first time in January 1959 from a female adult mosquito of the Culex neavei species in the area of Natal, South Africa (McIntosh 1985), a location equivalent to a tropical coastal lowland and savannah woodland, and was named after a river in Swaziland (Woodall 1964).
USUV is a flavivirus (Flaviviridae family) belonging to the Japanese encephalitis serocomplex (De Madrid and Porterfield 1974, Poidinger et al. 1996). The USUV genome is a positive-sense, single-stranded RNA genome of 11,064–11,066 nucleotides length (Weissenbock et al. 2002, Bakonyi et al. 2004, 2007) with a type I cap structure and no poly(A) tail (Hurrelbrink and McMinn 2003). It is organized as a typical flavivirus with one open-reading frame encoding a 3434-amino-acid-residue polyprotein, which is subsequently cleaved into three structural (core, membrane, and envelope) and eight nonstructural (NS1, NS2A, NS2B, NS3, NS4A, 2K, NS4B, and NS5) proteins (Bakonyi et al. 2004).
Phylogenetic analysis of the NS3 and NS5 coding regions as well as full-length genome nucleotide sequences of the USUV reference strain from South Africa (AY453412) allowed its classification within the mosquito-borne cluster of flaviviruses, close to Murray Valley Encephalitis virus and Japanese Encephalitis virus (Fig. 1) (Kuno et al. 1998, Cook and Holmes 2006). At present, when comparing the three USUV full-length genome nucleotide sequences from South Africa (AY453412), Vienna (AY453411), and Budapest (EF206350), they exhibit 97%–99.9% and 99% homology for nucleotides and amino acids, respectively (Bakonyi et al. 2004, 2007). Further, the partial sequences of the E protein coding region of 12 strains isolated in Austria show 95.9%–96.2% nucleotide identity and 98.9%–99.1% amino acid identity to the reference strain (Chvala et al. 2007).
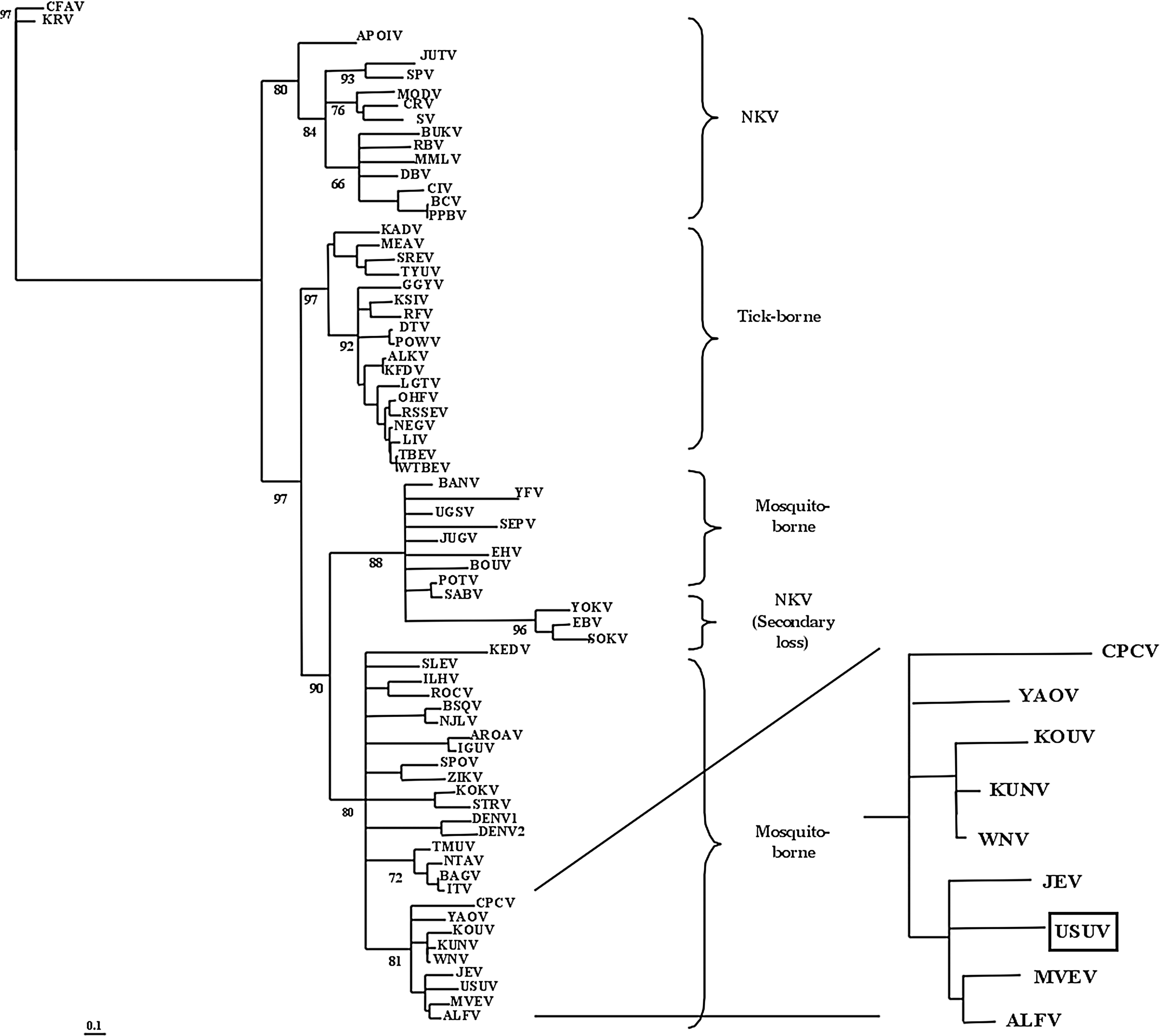
Localization of Usutu virus (USUV) within the mosquito-borne cluster of the flaviviruses. The analysis of the NS5 gene allowed the localization of USUV close to Japanese encephalitis virus (JEV) and Murray Valley encephalitis virus (MVEV), together with Cacipacore virus (CPCV), Yaounde virus (YAOV), Koutango virus (KOUV), Kunjin virus (KUNV), West Nile virus (WNV), and Alfuy virus (ALFV) (with permission from Cook and Holmes 2006).
Although no sequence data from USUV isolates from Africa besides South Africa are available, one subtype isolated from Culex perfuscus was serologically identified in the Central African Republic in April 1969 (Institut Pasteur de Bangui 1969). A year earlier and the following years, another subtype was reported in Yaounde (Cameroun), but it was later recognized as a different viral species named Yaounde virus in the light of further serological characterization (CDC 1985, CRORA 2005).
The biological characteristics of USUV have been analyzed by demonstrating its infectivity for several continuous and primary cell lines derived from various animal species and human [i.e., Vero, ED, MDBK, PK-15, RK-13, MDCK, DK, CR, BHK-21, BF, C6, TH1, EqK, GEF, LLC-MK2, HeLa (Bakonyi et al. 2005), and PS (De Madrid and Porterfield 1969, Stim and Henderson 1969)].
Geographical Distribution
After its first isolation in South Africa, USUV was isolated in Senegal, Central African Republic, Burkina Faso, Cote d'Ivoire, Nigeria, and Uganda (CRORA 2005) [Institut Pasteur de Bangui (Digoutte 1969), CRORA (Robin 1975, Digoutte 1995, 1998)] and USUV antibodies have been detected in wild birds in Morocco (Figuerola et al. 2009). USUV was reported in several European countries as listed earlier (Fig. 2). It is striking that USUV has been isolated only in African countries in which entomological surveillance programs have been undertaken, suggesting that its geographic distribution may be much wider than the reported circulation. Therefore, it would be reasonable to investigate further USUV circulation in other African countries.
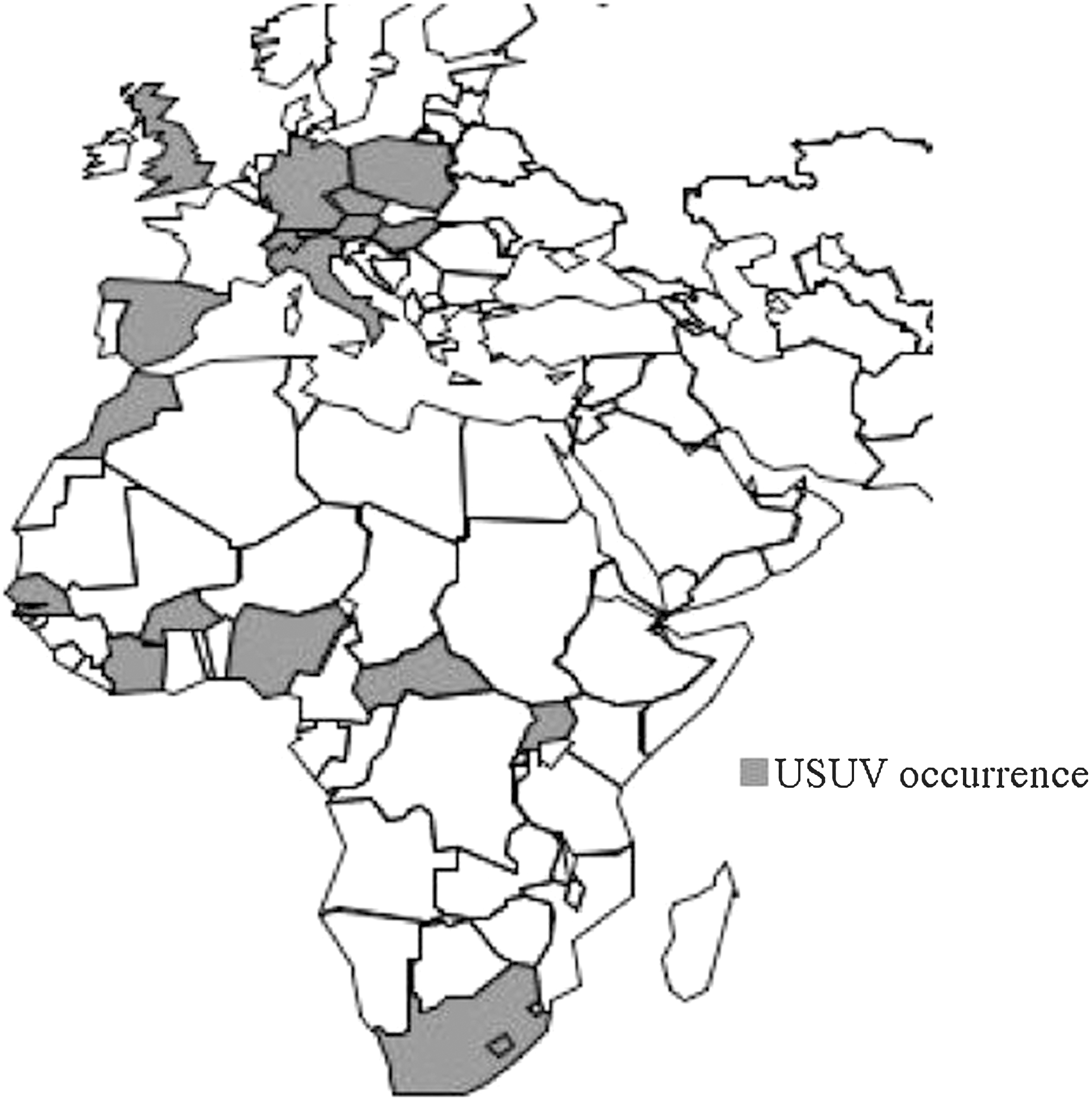
Geographical distribution of USUV. USUV has been detected in Europe, namely in Austria, Hungary, Switzerland, Italy, and Spain. Additionally, serological evidence of USUV circulation has been reported in Germany, the United Kingdom, Czech Republic, and Poland. USUV has been isolated in several African countries, such as South Africa, Senegal, Cote d'Ivoire, Burkina Faso, Nigeria, the Central African Republic, and Uganda. Further, seroprevalence in wild bird populations has been reported in Morocco (Weissenbock et al. 2002, Buckley et al. 2003, Bakonyi et al. 2007, Linke et al. 2007, Busquets et al. 2008, Hubalek et al. 2008a, b, Lelli et al. 2008, Figuerola et al. 2009, Pecorari et al. 2009, Steinmetz et al. 2011) (CRORA 2005).
In Senegal, where a continuous entomological surveillance program of the Institut Pasteur de Dakar started in 1972, USUV has been repeatedly isolated, providing information at the country level in Africa. The first USUV isolation occurred in June 1974 in Kedougou, followed by further isolations in November/December 1993 in the area of Barkedji and in July 1998 again in Kedougou (Fig. 3) [CRORA (Robin 1975, Digoutte 1995, 1998)]. Until 2003, only seven strains had been isolated with minimum infection rates (i.e., number of USUV isolates/number of mosquitoes per species collected) of 0.06%–1.35%, and therefore, the low level of circulation of USUV attracted only little attention. A sudden increase of USUV isolations was noticed in the year 2003, as 141 strains were isolated from Cx. neavei species in the area of Barkedji between October and December, with a higher minimum infection rate of 2.31% (CRORA, unpublished data). In Senegal, USUV has been mainly isolated from Cx. neavei but also from Cx. perfuscus and Aedes minutus mosquitoes. Isolations were restricted to the rainy season (CRORA 2005) (CRORA, unpublished data). The reason for the higher isolation number in 2003 remains unclear, but it is noteworthy that, at the same period, USUV was isolated in the Cote d'Ivoire, Burkina Faso, and some countries in Europe, suggesting a circulation at the regional level with potential interaction with Europe. Phylogenetic analysis of isolates in Senegal and other regions where USUV circulation has been reported should shed light on this question.
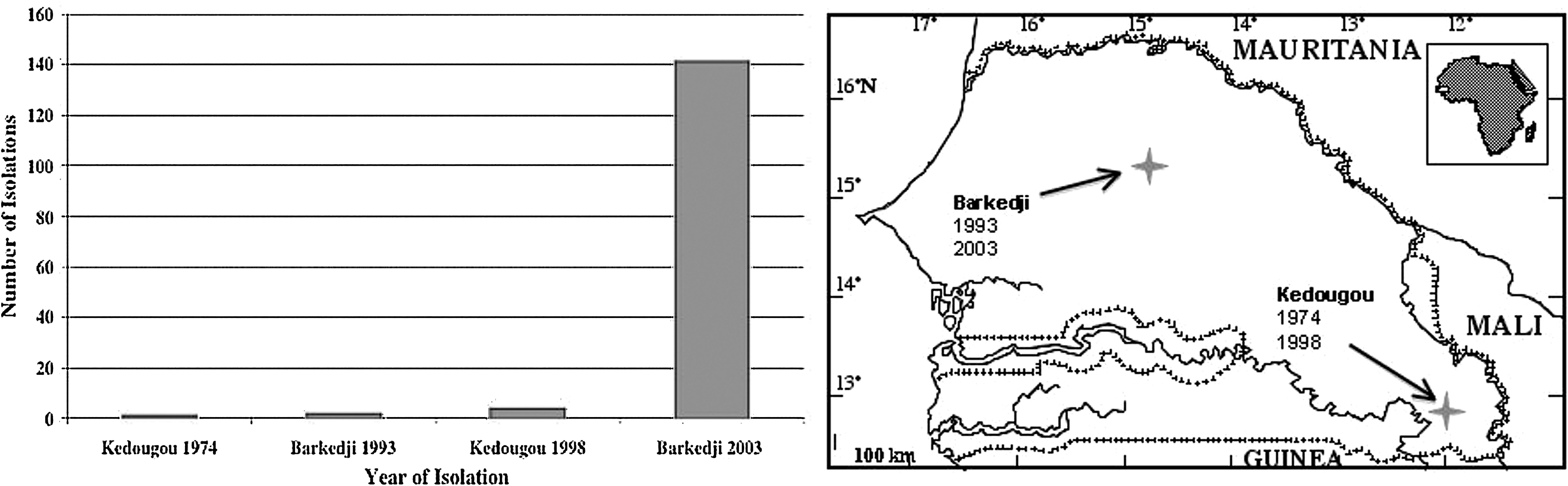
USUV isolations in Senegal. In Senegal, USUV isolation has been reported from two areas, Kedougou and Barkedji. USUV has been isolated for the first time in Senegal in the year 1974, followed by two isolations in 1993, and four isolations in 1998. In the year 2003, a total number of 141 isolations have been reported [CRORA (Robin 1975, Digoutte 1995, 1998, CRORA, unpublished data)].
Host Range of USUV in Africa
USUV has been mainly isolated from mosquitoes of the Culex species, twice from human serum, and four times from bird species (Table 1) (CDC 1985, CRORA 2005, CRORA, unpublished data).
Mosquitoes
Mosquitoes of the genus Culex are found in all zoogeographic regions. Several species are involved in transmitting Japanese encephalitis in Orient, encephalitis viruses in North America, and also Murray Valley Encephalitis virus and Ross River virus in Australia. Culex mosquitoes are known as vectors for arboviruses such as WNV, which is transmitted by three closely related species: Cx. univittatus, Cx. neavei, and Cx. perexiguus. Given the close morphological characteristics between Cx. univittatus and Cx. neavei, many reported isolations of USUV from Cx. univittatus might also actually be from Cx. neavei and vice versa (Jupp 1971). In general, mosquitoes of the Culex genus are considered as ground pool breeders, but larvae have been also found in leaf axils, tree-holes, and crab holes. The genus contains mostly ornithophilic species (Clements 1999); however, many species have been reported to have a large host range and feed not only on birds but also on amphibians, reptiles, humans, and other mammals (Chandler et al. 1975, 1976, Ba et al. 2006) (Walter Reed Biosystematics Unit 2010). Culex females usually bite at night and prefer the canopy (Lundström 1996, Anderson et al. 2004) where, in accordance with their roost and nest behavior, a high abundance of birds is observed. Experimental infection of Cx. neavei with USUV demonstrated that 2 of 10 mosquitoes were found positive at 14 days postoral infection, although the virus transmission to hamsters failed (McIntosh 1985). Additionally, a few isolations were reported from the mosquito species Cx. perfuscus and Cx. quinquefasciatus. Regarding its bionomic and ecology, the latter might be of great interest in the USUV transmission cycle. Cx. quinquefasciatus belongs to the Cx. pipiens complex, which has been shown to be involved in USUV epidemic transmission in Europe (Brugger and Rubel 2009). In addition, Cx. quinquefasciatus is the most present in the domestic environment and the most anthropophilic among the known USUV vectors. Further, the species has a population dynamic not related to rainfall, colonizing artificial habitats such as polluted water in artificial containers or edifices as ditches, catches, and basins, because of its steady presence in domestic environment. USUV isolations have been also reported from mosquito species of the genus Aedes, Mansonia, and Coquillettidia (CRORA 2005) (Williams et al. 1964). Like Culex mosquitoes, Coquillettidia mosquitoes are mainly bird-biting species, whereas Mansonia africana, which is confined to Africa, is mainly attracted to mammals and humans (Walter Reed Biosystematics Unit 2010).
Considering the bionomics of different mosquito species and their association with USUV in nature, Cx. neavei may play a key role in the enzootic transmission of USUV in Senegal, whereas species such as Cx. quinquefasciatus may represent a bridge or an epizootic–epidemic vector.
Human
So far, in Africa, only two isolations of USUV from human serum have been reported, with very limited information and clinical description. The first case occurred in 1981 in the Central African Republic and was a patient with fever and rash (Institut Pasteur de Dakar 1982–1984), and the second case was a 10-year-old patient with fever and jaundice identified in Burkina Faso in 2004 (CRORA, unpublished data). These cases show that USUV can infect humans and play a role as a human pathogen, occurring with benign infection symptoms such as fever and rash but also jaundice. As exemplified by WNV or CHIKV, more virulent strains can emerge and constitute a serious threat to human health, as the observed USUV neuroinvasiveness in an immunocompromised patient in Italy (Pecorari et al. 2009). However, the number of cases of human USUV infections is rather limited and more clinical work on infected patients is necessary to define its pathogenicity for humans.
Rodents
The isolation of USUV has been described from a rodent species (Praomys) in the Central African Republic, but it turned out to be the serologic variant S-t Y 276, which was later named Yaounde virus. Even though no isolation from mammals has been reported, guinea pigs and newborn and weaning mice have been shown to be sensitive to intracerebral inoculation with USUV (CDC 1985). A rodent species (Arvicanthis abyssinicus) showed a trace of viremia after intraperitonial inoculation (Simpson and Lule 1964). However, the available information is not enough to draw a conclusion about rodents serving as hosts for USUV, suggesting that fieldwork is needed to identify potential rodent hosts.
Birds
USUV has been isolated from the bird species piping hornbill (Bycanistes sharpei), little greenbull (Andropadus virens), and Kurrichane thrush (Turdus libonyanus) in Nigeria in 1972 (CDC 1985). Additionally, antibodies against USUV were detected in a blackbird (T. merula) in Morocco (Figuerola et al. 2009). This is consistent with the fact that many mosquitoes of the Culex subgenus are ornithophilic and allows first speculations about USUV transmission cycles. However, all mentioned birds are not migratory species, suggesting that they are probably not involved in dispersing the virus to Europe. In 2005, serological studies demonstrated the presence of USUV antibodies in numerous migratory wild birds in Austria, such as the kestrel (Falco tinnunculus), the reed warbler (Acrocephalus scirpaceus), the marsh harrier (Circus aeruginosus), the barn-swallow (Hirundo rustica), the lesser whitethroat (Sylvia curruca), the whitethroat (Sylvia communis), the garden warbler (Sylvia borin), the house martin (Delichon urbica), and the pied flycatcher (Ficedula hypoleuca) (Meister et al. 2008). These species might be candidates for the introduction of USUV into Europe but remain to be further investigated.
Prospects for Future Research in Africa
Besides several reported isolations, there are very limited data available about the host range, phylogeny, and ecology of USUV in Africa. Given its origin in that continent, it is essential to gain a better knowledge about this area to understand the evolution of USUV and its emergence in Europe. The first cases of human infections, already demonstrating its potential severity as a human pathogen, occurred in Africa before the first European cases in 2009 in Italy (Cavrini et al. 2009, Pecorari et al. 2009). The similarities of its pattern with the WNV scenario, the neuroinvasiveness in immunocompromised patients, as well as its experimental neuropathogenicity in mice (Weissenbock et al. 2004) emphasize the need to be cautious about its potential threat to human health. Many arboviral infections have been observed to either be asymptomatic or occur with an influenza-like illness. Nevertheless, the mosquito-borne flaviviruses also contain important human pathogens that cause central nervous system disease, coma, or death. In this light, there is an urgent need to understand the underlying mechanisms of flaviviruses emergence, and USUV might be a useful model as it emerged in several European countries within a limited time period and was able to adapt to new hosts and environments (Weissenbock et al. 2002, Buckley et al. 2003, 2006, Bakonyi et al. 2007, Lelli et al. 2008). Reported infections of humans indicate a role of USUV as human pathogen; however, more clinical research is needed to estimate the risk for human health. The investigation for the seroconversion for USUV in human populations living in concerned areas might give a better idea about its burden as a human pathogen. However, as the observed symptoms of human USUV infections are not very specific, it will also be meaningful to develop better diagnostic tools taking into account USUV variability in Africa and enabling an easier detection of USUV infections, such as the recently developed RT-PCR assay for the identification of USUV in human plasma, serum, and cerebrospinal fluid (Cavrini et al. 2011).
To date, transmission cycles of USUV in Africa are unclear, as the available information about the primary vectors, vertebrate hosts, and the ecology of the virus are very scarce. The recent observations of USUV in Africa demonstrate that the virus is isolated mainly from Culex mosquitoes (CRORA 2005), but the vector competence of the different mosquito species found infected needs to be investigated and combined with their feeding habits to identify potential vertebrate hosts for USUV, to gain further insight into its transmission cycle in nature.
As mentioned earlier, USUV has been isolated from nonmigratory birds in Africa. However, seroprevalence studies in Europe (Meister et al. 2008) identified several bird species possibly involved in virus dispersal, but it has to be confirmed by investigating bird populations in areas where USUV has been isolated in Africa.
Additionally, the phylogenetic analysis of USUV strains might help to understand the dispersal of the virus within Africa and Europe and to gain insight into the interactions between these two areas. At present, only three full-genome sequences of USUV strains are available: the reference strain from South Africa (AY453412) as well as the strains that emerged in Austria (AY453411) and Hungary (EF206350). Therefore, further sequence analysis of isolates from Africa is needed.
In conclusion, to understand the full range of virus characteristics and factors of emergence, it is necessary to not only follow the development of its emergence in Europe but also understand the relationship to the original strains that have been circulating in Africa for a longer period. Any information obtained about their characteristics, transmission dynamics, and impact on host ecology would be a valuable asset to understand the transmission dynamics in Europe, the dispersal, and ways of adaptation of USUV.
Footnotes
Acknowledgments
B. Nikolay is awardee of a scholarship by the Austrian Federal Ministry for Science and Research (BMWF). The authors thank the two anonymous reviewers for their comments that helped to improve the paper.
Disclosure Statement
No competing financial interests exist.