
Abstract
Investigating the prevalence of anti-influenza A viruses (AIV) antibodies in wild birds can provide important information for the understanding of bird exposure to AIV, as well as for prevention purposes. We investigated AIV exposure in nature by measuring the prevalence of anti-AIV antibodies in the nests and adults of an abundant and anthropophilic waterbird species common around the Mediterranean sea, the yellow-legged gull (Larus michahellis). Sampling took place in two colonies located in the gulf of Gabès in southern Tunisia: Sfax and Djerba. Antibodies were detected in the two sites, with higher prevalence in adults, eggs, and nests at Sfax than Djerba. Across both colonies, clutches that were laid later in the season, and, thus, more likely by younger parents, showed lower prevalence. Using patch occupancy modeling applied to egg clutches, we found that it is unnecessary to sample all the eggs in a given nest; nest status (antibody positive or negative) can be reliably estimated from a single egg. Differences in the density of birds, notably Larids, between the two sites may explain the observed differences in prevalence. The higher concentration of Larids in the Sfax colony could favor the transmission of AIV to yellow-legged gulls. This study highlights the importance of further developing ecological-based approaches to the factors determining the circulation of infectious agents in species such as the yellow-legged gull, which exist at the interface between diverse biological communities and human activities.
Introduction
The antibody quantification approach is especially useful when the proportion of individuals shedding virus at any point in time is low, which is common in the case of avian influenza (e.g., detection of viruses in only 0.5% of more than 300 yellow-legged gulls) (Larus michahellis) droppings sampled; (Lebarbenchon et al. 2010). Thus, when used on long-lived birds, the antibody detection approach enables one to detect antibodies that may have persisted after exposure to a virus several years earlier. This method provides one complementary tool for monitoring AIV in the wild, although limited information, concerning where and when the infection occurred, could be provided. Further, the presence of antibodies in egg yolks can reflect the past exposure of mothers to infectious agents without the need to capture adults (Gasparini et al. 2001, Grindstaff et al. 2003, Trampel et al. 2006, Boulinier and Staszewski 2008, Pearce-Duvet et al. 2009). Egg sampling presents some important advantages, namely that it provides one possible way for getting large samples from birds that are not easy to trap and sample. It also presents some limitations, in that it can only be applied for nonprotected species whose eggs can be readily collected without restrictions. Moreover, since detection of antibodies in the eggs means that their mother has somewhere been in contact with the virus during its life, this method also provides only limited information concerning the dynamics of virus transmission in the wild.
In this article, we used data on anti-AIV antibodies in the plasma and eggs of yellow-legged gulls (L. michahellis) breeding in two distinct sites in south-eastern Tunisia to investigate small-scale spatial variation in bird exposure to AIV. This species nests in large colonies and is a predator and scavenger, both behaviors that could enhance disease transmission. Moreover, since it is a long-lived bird, the yellow-legged gull offers a suitable model for investigating bird exposure to pathogens through antibody detection. Since the gulf of Gabès is at a migratory crossroad for palearctic birds moving between two epizootic areas of AIV, sub-Saharan Africa and Western Europe (Olsen et al. 2006, Gaidet et al. 2007, Jourdain et al. 2007), and given its importance as an overwintering area (Van dijk et al. 1986, Isenmann et al. 2005), it may be one of the most important high-risk zones in the Mediterranean; anti-AIV antibody prevalence in waterbirds breeding in this area is, thus, expected to be high overall. In particular, anti-AIV prevalence should be highest in the Sfax salina, as it is the most attractive site for waterbirds within this gulf and is, thus, expected to be a hotspot of bird exposure.
We also investigated factors affecting the detection and occurrence of AIV antibodies in yellow-legged gull nests. In doing so, we considered eggs from the same nest as sampling replicates of the same subject (i.e., the nest) and used a capture-recapture-like approach specifically conceived to estimate site occupancy rate by a given species from count replicates while accounting for the effects of various sampling and site parameters on both detection and occurrence probabilities (see Mackenzie et al. 2002, 2003). The use of this estimation method in the context of our study allowed us to check whether the probability of detection of anti-AIV antibodies in a given nest, and, hence, whether the deduced nest status, varied with the laying order of the sampled egg. We expected the probability of detecting antibodies in a given nest to vary with the laying rank of the sampled egg because of possible differences in antibody quantity among eggs of the same clutch, due to earlier suggestions of differential antibody transmission rates from the mother to the eggs reported in other bird species (Groothuis et al. 2006, Hargitai et al. 2006). This method also offered the possibility of estimating the prevalence of anti-AIV antibodies in yellow-legged gull nests, while controlling for imperfect antibody detection, and to check whether antibody occurrence probability varied among nests according to laying date and to breeding site. Given the fact that the laying date of females is likely related to their ages (older ones laying earlier) (Sydeman et al. 1991) and could, thus, be related to their history of risk of exposure to virus, we expected the prevalence of antibodies to be higher in early nests compared with late ones.
Materials and Methods
Study species
The yellow-legged gull is a colonial seabird of the family Laridae. It is long lived and shows strong interannual breeding site fidelity (Cramp and Simons 1983). In the studied area, egg laying occurs between late March and mid-April, with clutch size typically ranging from 1 to 3 eggs. As in other gull species (Sydeman et al. 1991), individuals that lay earlier in the season are often older/more experienced than individuals that lay later. In the Mediterranean region, the yellow-legged gull is a common and widespread species, where it is locally regarded as a pest and subject to population control measures (Vidal et al. 1998, Bosch et al. 2000, Oro and Martinez-Abrain 2007). This species is an opportunistic feeder; it not only forages at sea but also predates on other birds, eats earthworms, and feeds on refuse dumps (Ramos et al. 2009). Since this bird is anthropophilic, often nesting in urban areas and feeding on trash dumps, it may be more likely to exchange pathogens with humans and domestic birds (Bonnedahl et al. 2009). Further, its wide distribution and its feeding behavior make it an important species for monitoring the environment (Abdennadher et al. 2010).
Study area
Egg and blood sampling took place during the period between March 25 and April 11 2009 in two yellow-legged gull breeding sites situated 100 km apart in the gulf of Gabès, in southeastern Tunisia (Fig. 1). The yellow-legged gull is a common and abundant resident in this area, where it is often seen feeding on open air rubbish dumps and discards of commercial fisheries. The sampled colonies are located on opposite sides of the gulf: Sfax salina to the north (34°42′28′′N–10°45′02′′E) and Djerba island (33°39′10′′N–10°58′59′′E) to the south (Fig. 1). Sfax salina extends 15 km south from Sfax city, along the Mediterranean coast, and is bordered on the west by Sfax suburbs. It is formed by more than 200 ponds interspersed with artificial silty dykes that are used as breeding sites by a great diversity of waterbird species (Chokri et al. 2008). The Djerba site consists of a small islet (of approximately 50 ha) situated between Djerba island and Zarzis peninsula.
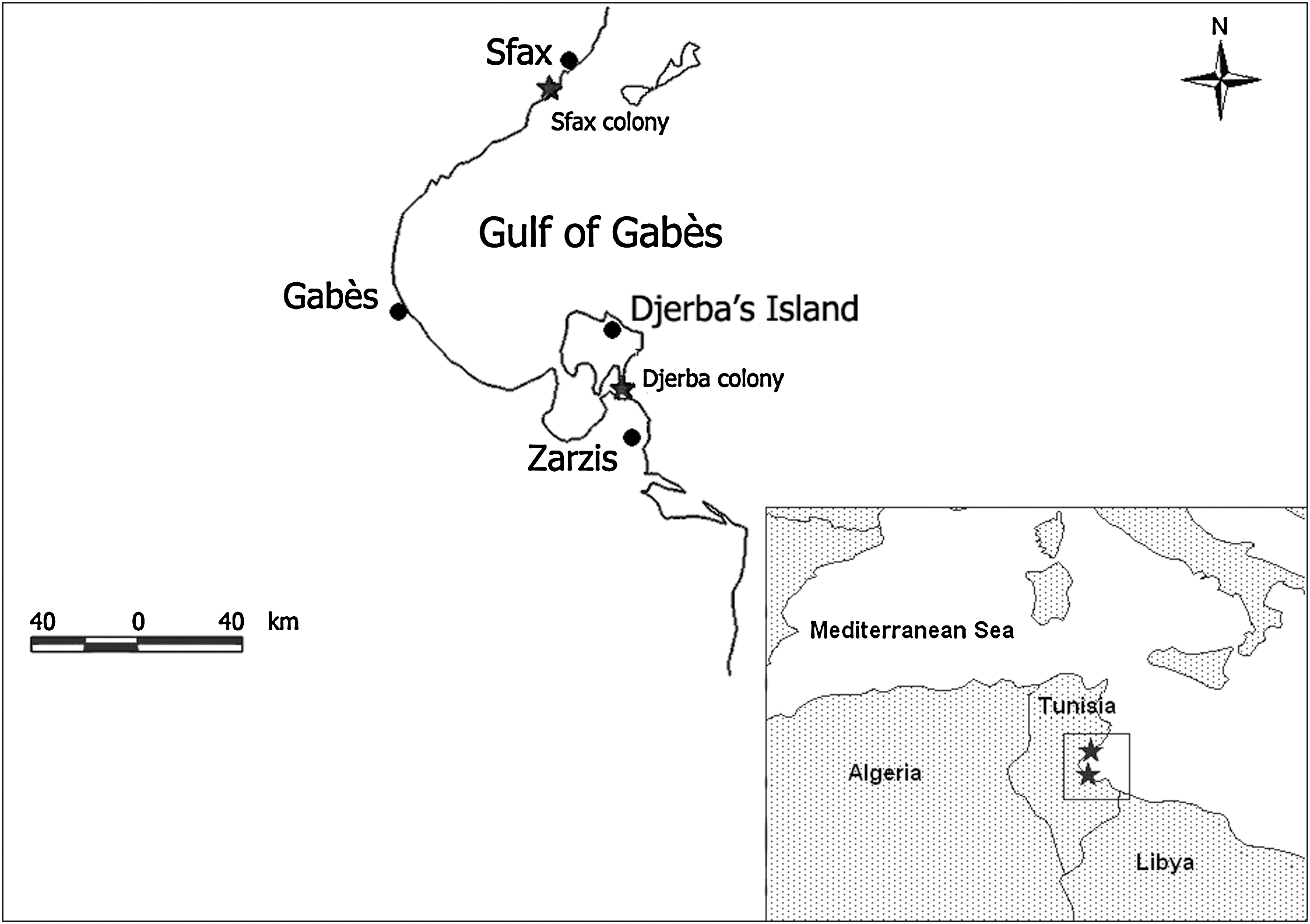
Map of the gulf of Gabès showing the location of the two studied colonies.
Data collection
Egg and blood sampling
To estimate the prevalence of anti-AIV antibodies in the studied colonies, we sampled both adults and entire clutches of a given group of nests. In each study colony, randomly selected nests were marked with a small wooden pole placed nearby. The nests were then checked every 1–2 days for collecting the newly laid egg(s), so that the entire clutch of each monitored nest was collected. Each egg was marked according to its rank and laying date and then replaced by a dummy egg to prevent females from abandoning their nests. The eggs were brought to the laboratory on the day of collection for processing. Collected eggs were, thus, 1 to 2 days old, and their yolk composition was not yet affected by embryonic development. The yolk of each egg was separated from the albumen, homogenized, and frozen at −20°C.
After clutch completion, we attempted to trap incubating parents by means of noose-carpet traps that we placed on the nests. Each captured bird was marked with a patch of paint on the head to avoid resampling. On capture, a 1-mL blood sample was taken from the brachial vein using a sterile syringe. We measured the head, bill, wing, and tarsus length (±0.1 mm) by using a caliper. We also determined body mass (±20 g) with a spring scale (PESOLA®) before releasing the bird. The blood sample was immediately transferred to heparinized tubes and maintained in a cooler while in the field (4°C). Once in the laboratory, it was centrifuged at 2500 rpm for 15 min. The plasma and blood cells were separately frozen at −20°C until immunological analyses could be performed. The sampled birds were sexed following the protocol described by Griffiths et al. (1998). DNA was extracted from blood using a DNeasy Blood & Tissue Handbook 07/2007 kit and used in PCR with primers 2550 Forward and 2718 Reverse to amplify introns from the CHD-Z and CHD-W genes (Fridolfsson and Ellegren 1999).
Immunological analyses
Plasma samples were directly used in the immunological assays. However, yolk antibodies first had to be extracted (Mohammed et al. 1986, Gasparini et al. 2001). Egg yolks were thawed and homogenized; a subsample of 800 mg was then taken. The yolk subsample was diluted 1:1 in phosphate-buffered saline, and the solution was vortexed after adding glass beads. An equal volume of reagent-grade chloroform was added to the mixture. The yolk chloroform mixture was then centrifuged at 16,000 rpm for 15 min, and the clear supernatant was used in the immunological assays.
We detected anti-AIV antibodies in gull plasma and yolk by using a commercial competitive enzyme-linked immunosorbent assay (ELISA) developed for use in birds (ID Screen® Antibody Influenza A Competition, IDVET). The assay is designed to detect antibodies directed against the internal nucleocapsid of the AIV, and, thus, it will pick up all AIV subtypes. Following kit manufacturer's instructions, the plasma and egg yolk samples were diluted 1:100 and then incubated at 37°C for 1 h. After a washing step, a conjugate solution was added to each well and incubated for 30 min at 21°C. The plates were then washed again; the substrate solution was added to each well and was incubated for 10 min at 21°C in the dark. Finally, a stop solution was added to each well to stop the reaction. Optical density (OD) was read at 450 nm by using an ELISA plate reader. A subset of samples was tested several times, both within and across plates. Repeatability tests showed that OD measurements were highly repeatable within (r=0.913, F [29,30]= 22.84, p<0.0001) and across (r=0.85, F [33,24]= 9.56, p<0.0001) plates.
According to the kit instructions, the results were expressed as the percentage competition (PC) between the OD of the sample being tested and the mean OD of a negative control (NC) sample, such that PC=(ODspecimen/ODNC)×100. The PC values were then transformed into percentage inhibition values (PI) by using the formula: PI=100 − PC. According to kit instructions, samples with a PI greater than or equal to 50% are considered positive, those with a PI less than or equal to 45% are considered negative, and those with a PI between 45% and 50% are considered doubtful. The same threshold of positivity was applied to the yolks, although the PI obtained for the eggs cannot be directly compared (in absolute terms) with those of the plasma, as the ELISA was developed for bird plasma. However, we verified that there was a strong correlation between the values obtained for eggs and that for the plasma of the female that laid these eggs (Hammouda et al. unpublished results).
Data analyses
First, plasma data were used to estimate the overall prevalence of anti-AIV antibodies in the study population (the proportion of positive birds relative to the total number sampled). We then compared antibody prevalence between the two colonies (Sfax vs. Djerba) and between males and females by means of χ 2 tests. Within breeding pairs for which both adults were sampled, we tested whether male and female serological status was related. This was done by using a Fisher exact test (appropriate for small sample sizes). All statistical tests were carried out using SAS software (SAS 1998).
We used yolk data to estimate the occurrence of anti-AIV antibodies in the sampled nests. Our sample units were not the individual eggs but rather the nests. Each nest was considered a sampling unit, and eggs from the same nest were considered sampling replicates of the same subject (i.e., the nest). We considered a nest positive if it contained at least one positive egg. We also assumed that negative eggs in a given nest were either true negative (absence of antibodies) or false negative (failure of detection). A negative egg (be it true or false negative) in a positive nest was considered a failure of detection of antibodies in that nest. We, thus, used the patterns of detection/nondetection of antibodies in the sampled nests to estimate the probabilities of antibody detection and occurrence in the nest population, using the capture-recapture-like methodology developed by Mackenzie et al. (2002, 2003) and implemented in program PRESENCE (available for download from
We employed a two-step process in our analyses. We first selected the best model for detectability by holding occurrence probability constant (ψ(.)) and evaluating two models: (1) one that assumed antibody detectability in a given nest was constant whatever the laying rank of the sampled egg (p(.)) and (2) one which assumed that antibody detectability varied according to the laying rank of the sampled egg (p(Rank)). In a second step, we used the best detectability model to estimate antibody occurrence probability. We defined a set of a priori models that we believed might explain antibody occurrence probability in our nests. The simplest model assumed that antibody occurrence probability was constant throughout the nests (ψ(.)). We then developed candidate models accounting for the possible effects of colony (Sfax vs. Djerba) and/or laying date. The competing models were ranked according to their Akaike's Information Criterion (AIC), and Akaike weights were applied to estimate the relative support of the models from the data (Burnham and Anderson 2002). To account for model selection uncertainty (Burnham and Anderson 2002, Wintle et al. 2003), we used a model averaging technique for estimating covariate effects (averaged β) and their 95% confidence intervals (CI) (Burnham and Anderson 2002). The relationship between a given covariate and antibody occurrence probability was significant if the estimated 95% CI of the corresponding averaged β included zero. We also computed odds ratios so as to interpret the effects of covariates on antibody occurrence.
Results
A total of 64 birds were captured and sampled across our two study colonies (Table 1). Overall anti-AIV antibody prevalence in plasma was 77%. It was not related to bird sex (females: 81%; males: 73%; χ 2 [1]=0.4830, p=0.4871). It did, however, differ significantly with location; prevalence was higher at Sfax (95%) than Djerba (68%) (χ 2 [1]=5.7676, p=0.0163). There was no association of antibody status within breeding pairs (Fisher's test: p=0.2727, n=11).
PI, percentage inhibition.
We sampled a total of 135 nests, corresponding to a total of 333 eggs (Table 1). Clutch size ranged from 1 to 3 eggs, with a modal clutch size of 3 eggs in both colonies (86% at Sfax and 53% at Djerba). The total number of positive eggs was 171, which corresponds to overall antibody prevalence in eggs of 51%: 66% at Sfax and 42% at Djerba (Table 1). Fifteen eggs had doubtful serostatus (Table 1) and were considered failures of detection (0) when modeling the probabilities of antibody detection and occurrence in the studied nests. The apparent antibody prevalence in the nests (i.e., the proportion of positive nests relative to the total number sampled) was 56% (71% at Sfax and 49% at Djerba). The percentage of positive nests whose all eggs were positive was 76%. Using multiple-raters Kappa test, we found a highly significant agreement level among eggs of the same clutch in the classification of the corresponding nest as positive or negative (Kappa [±SE]=0.82 [±0.06], Z=12.88, p<0.0001). The latter results show that within-clutch results were highly repeatable.
In our first step of capture-recapture model selection, we found that both detectability models showed a similar fit to the data (Model ψ(.),p(.): AIC=300.75, ΔAIC=0, weight=0.6759; Model ψ(.),p(Rank): AIC=302.22, ΔAIC=1.47, weight=0.3241). It did not matter if antibody detectability was assumed to be the same whatever the sampled egg or if it was assumed to vary with the rank of the sampled egg. In the constant detectability model, the overall detection probability of antibodies in the nests was 0.9015 (95% CI [0.8467–0.9381]). When egg rank was included as a sampling covariate, antibody detection probability tended to decrease with the rank of the sampled egg: 0.9198 (95% CI=[0.8341–0.9631]) according to egg 1, 0.9011 (95% CI=[0.8458–0.9380]) according to egg 2 and 0.8786 (95% CI=[0.7680–0.9405]) according to egg 3. However, the trend was not significant (β±SE=−0.23±0.32), which is consistent with the high within-clutch agreement that we found by using the multiple-raters Kappa test (please see above).
The constant detectability model was used for modeling antibody occurrence in nests. Our results gave less support to the constant occurrence model (ψ(.)), as it was the last ranked one (lowest weight) (Table 2). Conversely, the model including both colony and laying date as nest covariates was the most supported model, with an Akaike weight of 72% (Table 2). Using the model averaging technique, we found that the occurrence probability of antibodies in a given nest differed significantly between the two colonies and also according to laying date. The 95% CI associated with the β estimates of both variables did not overlap 0 (Table 3). The occurrence probability of antibodies in nests was almost thrice lower in Djerba compared with Sfax (odds ratio=0.36) and almost twice as high in early clutches compared with late ones (odds ratio=1.84). Overall, the estimated antibody prevalence in nests varied between 35% (late clutches in Djerba) and 79% (early clutches in Sfax) (Table 4).
K, number of model parameters; AIC, Akaike's information criterion; W, Akaike weight.
CI, confidence interval.
The estimates were obtained under model ψ(Colony,Laying date),p(.).
Discussion
We found a high prevalence of detectable antibodies against AIV in the two yellow-legged gull colonies we sampled, which underlines the potential importance of the yellow-legged gull in the epidemiology of AIV. Since gull species are relatively long lived (e.g., annual survival of adults >0.8), it is not unexpected that a high proportion of the population shows signs of having been exposed to AIV if anti-AIV antibodies persist over long periods in these species. Recent studies on related gull species living in different areas have also reported a high prevalence of anti-AIV antibodies (Brown et al. 2010, Velarde et al. 2010, Toennessen et al. 2011), which gives support to this hypothesis.
The yellow-legged gull nests in dense colonies and can feed on open air rubbish dumps. The aggregation of individuals at nesting and feeding sites is likely to lead to high rates of contact between individuals (McCallum et al. 2001), which could facilitate the transmission of AIV between individuals and lead to a high prevalence of viruses in the population (Bin Muzaffar et al. 2006). The fact that AIV are sensitive to elevated salinity (Stallknecht et al. 1990) and that the studied colonies are surrounded with saline water provides arguments supporting the idea that, in the study site, AIV epidemics may not be driven by water-borne transmission, but rather by direct bird-to-bird transmission. Further investigations of the aggregative behavior and interactions among birds at their feeding and nesting sites would tell us more about this issue (Munster and Fouchier 2009). Interestingly, our results show that antibody prevalence in the eggs of yellow-legged gulls nesting in the gulf of Gabès was higher than that reported for the same species nesting in the northern border of the Mediterranean (Pearce-Duvet et al. 2009). Long-term studies would need to be performed to test for the significance and meaning of this difference.
Our results also show that antibody prevalence did not differ between sexes. In the yellow-legged gull, males and females incubate eggs, rear offspring, and frequent the same feeding areas, and, thus, likely share the same risk of exposure to AIV. In contrast, antibody prevalence in plasma and also in nests was significantly higher in Sfax than in Djerba. This result could possibly be interpreted as a consequence of higher bird abundances, in particular those of Larid species, in Sfax salina compared with Djerba. Sfax salina is considered one of the most important bird areas in Africa (Fishpool and Evans 2001) and hosts the most rich and dense community of wintering and nesting waterbirds in Tunisia (Chokri et al. 2008). Over 30,000 birds belonging to 49 migrant species and 8,500 birds from 9 resident and breeding species are known to regularly inhabit this area and to share the same feeding and/or breeding sites every year (Chokri et al. 2008). However, in Djerba site, the yellow-legged gull is the unique breeding Larid species and shares the available breeding sites with only a small colony of little egrets that are not likely to represent a source of infection for gulls. We think that the higher density of Larids in Sfax would favor the transmission of Larid-related AIV subtypes to yellow-legged gulls, especially as they frequently prey on the chicks of the four other Larid species. However, we believe that investigating the prevalence of antibodies specific to H13 and H16 subtypes circulating in Larids is necessary for more firm conclusions. Further, it is to be underlined that alternative explanations of the difference in anti-AIV antibodies between the two sites could be proposed. For instance, it could be due to possible differences between the two studied colonies in terms of environmental or genotypic factors that affect the ability of birds to mount detectable immune response. In wild birds, phenotypic variation in the ability to mount immune response is known to have environmental and genetic components (Christe et al. 2000, De Neve et al. 2004). In particular, bird immune responsiveness can strongly be affected by food availability (e.g., Saino et al. 1997, Christe et al. 2000, De Neve et al. 2004). Further, evidence for family-related components in immune responsiveness in wild bird populations has also been found (e.g., Christe et al. 2000, Tella et al. 2000). Possible spatial variations in these environmental and genetic factors affecting the ability of birds to mount immune responses are, thus, supposed to result in similar patterns of spatial variation in antibody prevalence. This would imply that if the environmental conditions, notably the availability of food, permitted higher immune responsiveness in Sfax compared with Djerba, or if the yellow-legged gulls breeding in Sfax were genetically more able to mount immune responses against AIV than those breeding in Djerba, then the prevalence of anti-AIV antibodies would be higher in Sfax even if the exposure to AIV was the same in the two sites. Further investigations of yellow-legged gull immunocompetence in relation to habitat features, in particular to food availability, as well as to population density and genetic structure are likely to provide important information on this issue.
The capture-recapture approach we used allowed us to ensure that the detection probability of antibodies did not differ according to the laying rank of the sampled egg, although there was a tendency for decreasing detectability with increasing egg rank. This result would suggest that the sampling of only one egg per nest, of whatever rank, is efficient to provide unbiased results regarding the prevalence of antibodies in the nest population. Further, the approach we used allowed us to indirectly assess the relevance of female age as a predictor of the occurrence probability of anti-AIV antibodies in its nest. Our results support our prediction and would suggest that in the two studied colonies, the older the female (i.e., the earlier the laying date), the higher the occurrence probability of anti-AIV antibodies in her nest. Older females are likely to have been exposed to the infection for a longer time than the younger ones.
Mounting and maintaining an immune response is assumed to be energetically and nutritionally costly (Sheldon and Verhulst 1996). Consequently, to boost their immune system, birds have to reallocate some of their resources from other demanding functions, such as growth, development, molting, and reproduction (Ilmonen et al. 2000). The immune system may be adversely affected when such reallocation of energy or nutritional conditions are not ideal. In this context, AIV prevalence may be affected by different factors such as food supply, the age and the sex structure in a colony, and individual immunocompetence (Weber and Stilianakis 2007); and it should be possible to investigate such processes by monitoring the incidence of exposure in the young birds of the population.
In conclusion, our study suggests that the yellow-legged gull has the potential to play a major role in the epidemiology of AIV. It also suggests that even at the relatively small geographic scale of the studied area, there can be important variations in the level of exposure of individuals. We hypothesize that this spatial variation is related to differences in avifaunal density and diversity between the two studied breeding sites. Further investigations of the ecology of the yellow-legged gull and its interactions with the sympatric waterbird species in a series of comparable sites are needed to test our predictions.
Footnotes
Acknowledgments
This work was carried out as a part of the ARDIGRIP project supported by the Agence Inter-établissements de Recherche pour le Développement (France). Authorizations for working in the two studied sites and for bird and egg sampling were obtained from the forest service in the Tunisian Ministry of Agriculture (permit reference: 518–28/02/2009). Analyses were conducted at the PACE platform at CEFE-CNRS. Jessica Pearce-Duvet was a recipient of a Chateaubriand postdoctoral Fellowship, and the PhD of Audrey Arnal is funded by CNRS. The authors thank A. Kabaou, who helped a lot in data collection, and W. Alioua, who helped with the immunological analyses. They are also grateful to A. Karem (from the Forest service in the Ministry of Agriculture, Tunisia) and R. Machta (from the COTUSAL) for their helpful administrative support.
Disclosure Statement
No competing financial interests exist.