
Abstract
The aim of the study was to reveal new aspects of the role of flea vector taken from migratory birds by screening of specimens with molecular biological methods. A field study was done in fishponds in Slovakia. Actually, 47 fleas were collected from reed warblers (Acrocephalus scirpaceus) and their nests. DNA was extracted and analyzed for representatives of the orders Rickettsiales. A rickettsia that shares 99.7% of identity by gltA gene with Rickettsia africae was identified in Ceratophyllus garei collected from A. scirpaceus. Moreover, two Wolbachia sp. were also detected in fleas. This is the first record of R. africae and Wolbachia sp. identified so far in Central Europe in fleas collected from migratory bird returning from Africa. This molecular study extends the geographic range and vector spectrum of arthropod-borne agents.
Introduction
Birds have been neglected as potential reservoirs of rickettsioses. However, several studies showed the possibility of experimental infection of birds with Rickettsia rickettsii (Lundgren et al. 1966). Rickettsiae were found in ticks taken from birds in United States and indicated the presence of R. rickettsii and the Rickettsia sp. of the typhus group (Clifford et al. 1969). Sera obtained from ostriches on farms around Zimbabwe have antibodies to Rickettsia africae (Kelly et al. 1996). Several species of birds were found to be seropositive to Rickettsia sibirica (Hubalek 2004). Moreover, wild birds were recently described as an important reservoir of pathogens and disseminator hosts of arthropods (Santos-Silva et al. 2006, Spitalska et al. 2006), and Rickettsia bellii and Rickettsia parkeri were found in Amblyomma nodosum ticks collected from Passeriformes birds in the Atlantic forest, eastern Brazil (Ogrzewalska et al. 2009). Rickettsia helvetica was predominant among the polymerase chain reaction (PCR)-positive samples in ticks gathered from Passerine migratory birds in Sweden (Elfving et al. 2010). Three percent of bird blood samples were positive for Rickettsia spp. in Cyprus (Ioannou et al. 2009).
The role of birds in dissemination of ticks carrying the agent of Lyme disease, Borrelia burgdorferi, is well known and was described as an important component in the global epidemiology and evolution of Lyme disease (Duneau et al. 2008).
Birds are known to be colonized by a number of blood-sucking arthropods, including fleas, which hypothetically are able to transmit the variety of diseases. Several species of fleas (Siphonaptera) are notorious vectors of such devastating diseases as plague. Oriental rat flea Xenopsylla cheopis is the main vector of Rickettsia typhi, causative agent of murine typhus (Hackstadt 1996). However, according to T. Trilar et al. (1994), R. typhi may exist also in sylvatic flea Monopsyllus sciurorum sciurorum as well. Rickettsia felis is known to be associated with a cat flea, Ctenocephalides felis felis (Reif and Macaluso 2009). Moreover, a highly similar agent to R. felis, sequence of “RF2125”, was detected in a chicken flea, Echidnophaga gallinaceaa in Egypt, in a species that is capable of infesting a wide variety of birds and mammals (Loftis et al. 2006). A similar agent was identified in a human flea Pulex irritans in Hungary (Hornok et al. 2010).
Our interest was concentrated to a bird flea, Ceratophyllus garei, which is a holarctic species parasitizing almost exclusively birds and abundantly occurring in their nests (Beaucournu and Launay 1990). Blood sucking is an inevitable precondition for egg development in the female body. After a sufficient blood sucking flea leaves its host and lays eggs in a suitable substrate, females suck blood repeatedly before development of each subsequent clutch. It parasitizes in birds nesting directly on water tables, above it, or in its vicinity. Abundance of C. garei culminates in June and July, in synchronization with reproduction cycle of hosts (Cyprich and Krumpal 2001). This species was recorded in Slovakia as representatives of several genera of birds, for example, Anas, Aythya, Acrocephalus, Fulica, Larus, Ixobrychus, Motacilla, Tachybaptus (Kristofik et al. 2001, 2005, Cyprich and Krumpal 2006), and Panurus (Kristofik et al. 2007).
To the best of our knowledge, this flea species has not been earlier studied for the presence of rickettsial agents.
Materials and Methods
Fleas
The inspection of birds and their nests
Reed warblers (Acrocephalus scirpaceus) and their nests (112 in total) were studied in summer 2008 and 2009 at fishponds near Veľké Blahovo (48°03′05.39″N, 17°35′36.05″E) in Slovakia.
Collection of fleas
The birds were mist-netted and after taking the fleas, they were released. The fleas from the nests were extracted by means of the Tulgrens's funnels. Collected fleas were stored at 4°C until species determination and DNA extraction.
Identification of the species
The species of collected flea individuals was identified according to standard taxonomic keys (Rosicky 1957, Beaucournu and Launay 1990).
Molecular studies
Fleas were immersed in distilled water for 10 min, dried on sterile filter paper, and individually crushed with a sterile scalpel in Eppendorf tubes. Genomic DNA was extracted using QIAgen columns (QIAamp tissue kit; Qiagen, Hilden, Germany), according to the manufacturer's instructions. DNA from fleas were tested with the universal oligonucleotide primers fD1 and rp2, which amplify the main part of the 16S rRNA gene (Weisburg et al. 1989). Samples were also screened for the presence of rickettsial citrate synthase gene (gltA) using primers CS2d and CSEndr, which amplify a 1251-bp fragment of the gltA gene of Rickettsia, as previously reported (Roux et al. 1997). All primers were purchased at Eurogentec, Seraing, Belgium.
A negative control of DNA extracted from noninfected laboratory ticks and a positive control of Rickettsia conorii DNA were included in each test. Reactions were performed in automated DNA thermal cyclers (GeneAmp PCR System 2400 and 9700; Applied Biosystems, Courtaboeuf, France). The PCR products were verified by electrophoresis in agarose (1.5%) gel stained with ethidium bromide and examined under UV transillumination. The PCR products were purified using a QIAquick Spin PCR purification Kit (Qiagen) and sequenced using a DNA sequencing kit (dRhodamine Terminator cycle sequencing ready reaction; Applied Biosystems), according to the manufacturer's instructions. All sequences were assembled and edited with Auto Assembler software (version 1.4; Perkin-Elmer, Courtaboeuf, France)
Phylogenetic trees based on aligned complete sequences of the gltA gene were constructed by the NJ method. The numbers in the nodes represent bootstrap values. Sequences obtained were aligned and analyzed using the ClustalW and Mega v.4 programs.
Results
In total, we have collected 47 fleas. All fleas were identified as C. garei (Fig. 1). The DNA extracted from all 47 fleas was first tested in a standard PCR with universal eubacterial primers for 16S rRNA gene. All samples produced bands of different intensity. We have tried to directly sequence these amplicons; however, only in 6 cases, we obtained enough good-quality sequences to identify the bacteria. In all other cases, the sequencing presented multiple unreadable peaks. The results are presented in Table 1. Surprisingly, among evident environmental gamma-proteobacteria (vouchers CG02, CG03, and CG05), we have also identified representatives of obligate intracellular alpha-proteobacteria from Rickettsiales order, namely Rickettsia sp. and Wolbachia sp. The 997 bps-long sequence obtained from the flea voucher CG12 had 100% homology with Wolbachia sp. wRi (CP001391), yet uncultured obligate endosymbiotic bacterium from Drosophila simulans (Klasson et al. 2009). The voucher CG14 had 100% homology with Wolbachia pipientis strain EW-p. (Ryu et al. 2008), and the sequence was 960-bps long.

Photography of Ceratophyllus garei. The flea visualized in stereomicroscope SOLIGOR SZM-645 PV (magnification 40×), photograph done by Nikon Coolpix 4500.
The 172 bps sequence of rickettsial 16S rDNA (CG13) had 100% identity with multiple rickettsiae, including many pathogenic species. The discriminative power of the piece of 16S rRNA gene was not enough to identify the species level of rickettsiae. Since Rickettsia spp. plays an important role in human pathology, we have performed these results by amplification and subsequent sequencing of almost-complete genus-specific gltA gene. At the same time, we performed the screening by a standard PCR of DNA of all collected fleas.
We have identified rickettsial DNA in 15 of 47 (31.9%) fleas. We have succeeded in amplification of 1251 bps almost-complete gltA gene of Rickettsia sp. from C. garei in 2 of them (Genbank accession number HM538186). BLAST search showed that, surprisingly, it shares very high (99.7%) identity with R. africae, strain ESF-5 (CP001612) with only 3 single mutations (Genbank accession number is HM538186). The phylogenetic tree shows the position of Rickettsia from C. garei among recognized species (Fig. 2). Despite all our efforts, we could not amplify the rickettsial ompA gene from our samples (data not shown).
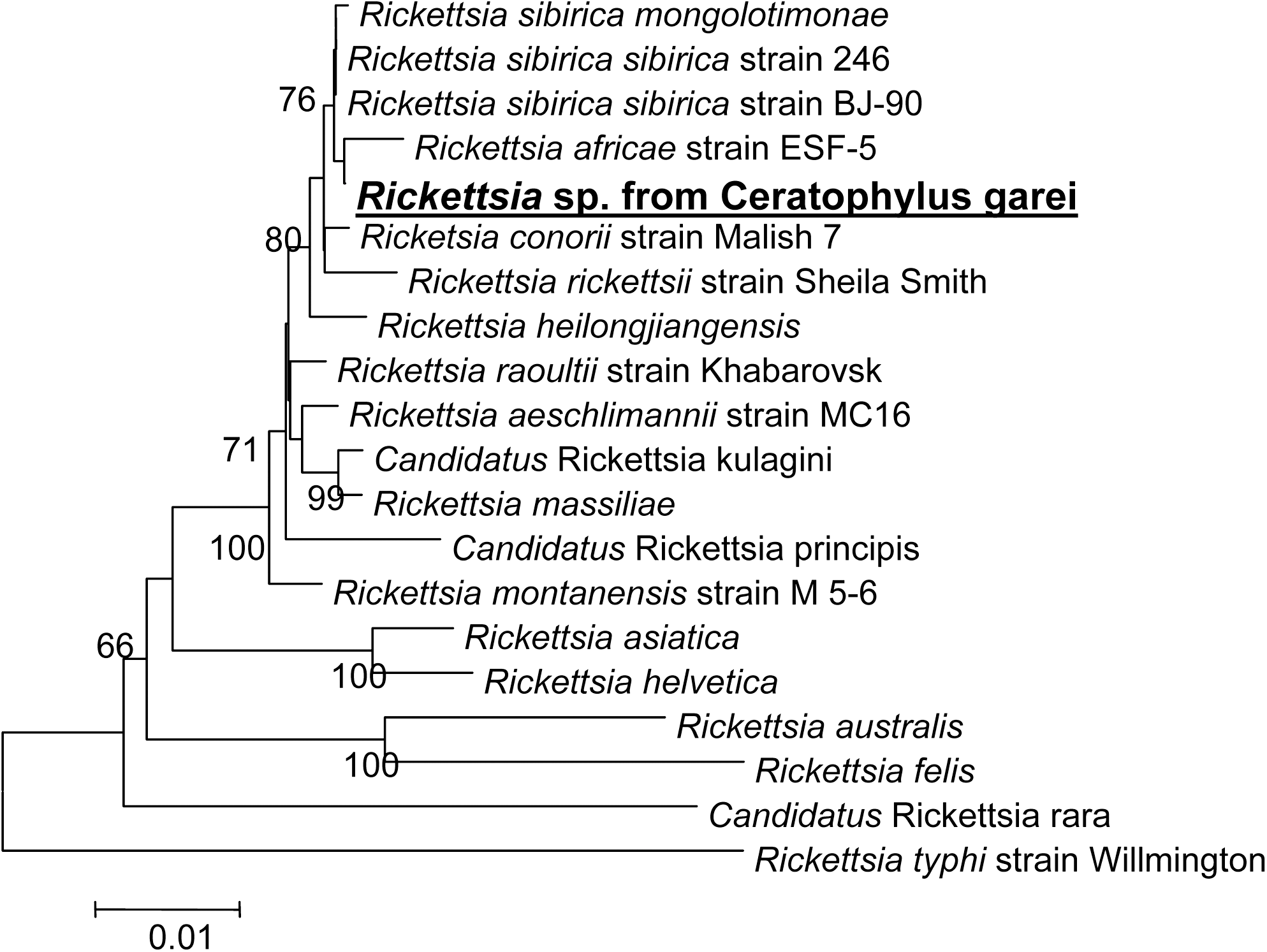
Phylogenetic tree of bacteria belonging to the genus Rickettsia inferred from the comparison of 16S rDNA sequences by the neighbor-joining method. The numbers at nodes are the bootstrap values obtained from 100 resamplings. The scale bar represents a 1% difference in nucleotide sequences.
Discussion
We provide a first evidence of the presence of Rickettsia sp. and Wolbachia sp. in C. garei flea collected after blood meal on reed warbler. Reed warbler is a small passerine migratory bird whose breeding area reaches from Morocco and Ireland through almost entire Europe to Southwestern China. It winters in savannas of sub-Saharan Africa, from Senegal to Ethiopia, and southerly toward Republic of South Africa. A part of reed warbler's populations migrates to wintering places in the southwestern direction through Morocco and Algeria, whereas another part migrates in the southeastern direction through Egypt (Cepak et al. 2008). In Slovakia, it breeds in reed and cattail stands around water tables. It arrives to breeding places from April to the first half of May. Most pairs breed once a breeding season, but some pairs even twice. The nests are constructed exclusively by females from materials of plant origin sometimes also fastened by spider fires. Females lay mostly 4 eggs, and incubation lasts for about 14 days. The chicks are fed by both parents, first of all by insects and other little invertebrates. The chicks fledge after 9–13 days. The autumnal migration runs mainly in August and September (Fig. 3). Transmission of fleas between individuals can run horizontally (among individuals), vertically (from parents to chicks), and birds can be also infested by them when moving on the ground or in vegetation.
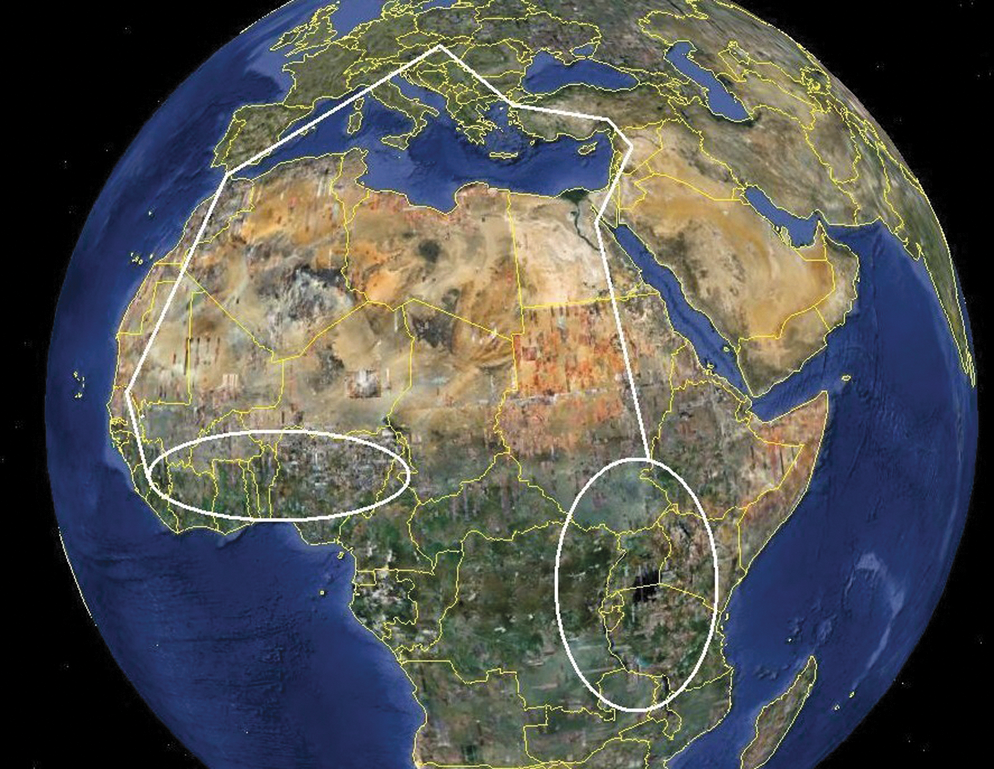
Migration flyways of Reed warbler. Flyways are marked by white lines.
Rickettsia identified in this study is genetically very close, although not identical, to a human pathogen, R. africae, and phylogenetically belongs to the same spotted fever group. We may hypothesize that the rickettsia in migratory bird fleas may be of the same origin as R. africae routinely find in African ticks.
R. africae, a causative agent of African tick-bite fever (ATBF), is preferably transmitted by Amblyomma ticks. The disease was described as an emerging, frequent rickettsiosis in the African continent (Cazorla et al. 2008, Jensenius et al. 2009). The ATBF in patients with a febrile rash after returning from endemic areas was illustrated as an indispensable, most frequent diagnostic conclusion (Jensenius et al. 2004, Jensenius et al. 2009). Given the current increase of international safari tourism to sub-Saharan Africa, several cases of sub-acute neuropathy after ATBF may be encountered in Europe and elsewhere in the years to come. However, no transmission of Rickettsia sp. in fleas captured from birds was ever described earlier; moreover, no spotted fever group rickettsia was reported in fleas.
We were also able to identify a bacterium either identical or closely related to yet uncultured Wolbachia sp. strain wR1 identified in D. simulans, as well as W. pipientis isolated from the gut of Drosophila.
Wolbachia is a genus of bacteria that infects different arthropod species, including insects. Several species of fleas (Siphonaptera) (Gorham et al. 2003, Dittmar and Whiting 2004), lice (Phthiraptera) (Covacin and Barker 2007), or ectoparasites of mammals and birds have been shown to harbor this germ. It is one of the world's most common parasitic microbes and is possibly the most common reproductive parasite in the biosphere. Recent identification of this microbe in the small pigeon louse has triggered a proposal of a novel mechanism for the horizontal transfer of Wolbachia between different species of lice from birds (Covacin and Barker 2007).
Screening related to bacterial (Bencina et al. 2003, Laukova et al. 2003) or viral infections (Ernek et al. 1973, Cernik et al. 1985, Literak et al. 2006) in territories of lakes is highly beneficial. Lakes are regions characterized by a high prevalence of migratory birds; thus, they represent a natural biotope of risky zones with possibilities to introduce various infections. Method of transmission, period of dormancy, ease of contagiousness, and relative danger can differ from one disease to the next (Santos-Silva et al. 2006). Blood-sucking arthropod vectors, ticks and fleas, and parasites of birds are able to transmit infectious agents between animals and/or humans, and are, therefore, crucial for epidemic studies.
Fleas are known to be intermittent feeders capable of multiple blood meals per generation, facilitating the efficient transmission of rickettsiae to several disparate hosts within urban/rural ecosystems (Gillespie et al. 2009). Birds infested with C. garei flea species were traced on its flyway to tundra in Russia (Bogdanov 1981), they were collected from sea bird nests in mainland Norway, or recorded as the only species in Larus gull's kittiwakes and shags nests in Spitzbergen (Mehl 1992). Nevertheless, no examination of infestation with Rickettsia sp. in the flea parasite of migratory birds was ever published earlier. Abundance of parasites and not a possibility of bacterial intervention were in the scope of studies provided with C. garei thus far.
This is a report of the presence of the spotted fever group rickettsia close to R. africae in Europe, the first detection of rickettsial agents in C. garei flea and the first detection of spotted fever group rickettsia in a flea. This finding evokes new insight into the ecology of rickettsiae in the context of possible ways of dispersion, and it extends our knowledge of the area of occurrence in vectors and the spectrum of hosts. Moreover, Wolbachia sp. was also detected in this flea species for the first time, which emphasizes the ubiquity of Wolbachiae in nature. Revision of the role of ectoparasites of birds in the maintenance and dissemination of rickettsia infection is still an open challenge.
Footnotes
Acknowledgments
This work was partly supported by grants VEGA No. 2/0016/08, 2/0031/11, 2/0071/09, and 2/0065/09 from the Slovak Academy of Sciences, Bratislava, Slovakia, as well as a bilateral project between CNRS France and SAS Slovakia, (No: 22525-CNRS, 2-SAS). The authors wish to thank RNDr. Ľubomír Vidlička CSc. from Institute of Zoology SAS, Bratislava, Slovakia, for a picture of C. garei.
Disclosure Statement
No competing financial interests exist.