
Abstract
Vaccinia virus (VACV) has been associated with several exanthematic outbreaks in bovine, human, and equine species in Brazilian rural areas. Little is known about VACV reservoirs, although it is believed that rodents could be associated with VACV outbreaks. With the goal of filling one more gap in the VACV ecological puzzle, the present work aimed at mimicking a potential transmission route of VACV between cows and rodents, both known as natural VACV hosts. Balb/c mice were exposed to feces of experimentally VACV infected cows for 20 days, and samples from these mice were examined by using molecular and serological tests. VACV DNA was detected in feces and blood samples after several days of exposure; infectious VACV particles were also detected in the feces. The presence of anti-VACV neutralizing antibodies in murine sera further suggested horizontal transmission. If the transmission model described here can be applied to natural environments, exposure to bovine feces could be considered a risk factor for the spread of VACV; consequently, the traditional use of bovine manure as a fertilizer in agricultural activities may be promoting the infection of rodents.
Introduction
Rodent species are considered potential OPV reservoirs and are associated with viral circulation during VACV outbreaks (Maiboroda 1982, Abrahão et al. 2009a). At least three different VACV strains were isolated in Brazilian forests in the past, away from rural environments: Cotia virus/SPAn 232 virus was isolated from sentinel mice in São Paulo state (Lopes et al. 1965); and BeAn 58058 virus was isolated from the blood of an Amazon rodent specimen belonging to the Oryzomys genus (Fonseca et al. 1998). In addition, biological and molecular studies demonstrated that the same VACV isolate circulated among rodents, bovines, and humans, during a VACV outbreak in Brazil (Abrahão et al. 2009a). Previous studies demonstrated that Balb/c mice intranasally inoculated with Brazilian VACV strains shed viral particles and DNA in their feces for at least 30 days postinfection. Sentinel-mice exposed to murine excrements contaminated with VACV also become infected and shed viral particles and DNA in their feces (Ferreira et al. 2008b). A similar viral shedding profile was shown for other OPV in rats (Maiboroda 1982). Moreover, long-lasting stability of VACV in murine feces exposed to environmental conditions was recently demonstrated, thus raising important questions related to viral circulation and maintenance among wild rodents (Abrahão et al. 2009c). In this line, preliminary studies carried out by our group demonstrated that lactating cows experimentally infected with VACV by scarification of their teats shed VACV in their feces for several weeks (Guedes et al. 2010). Although such results suggest that feces of infected bovines may represent a constant source of environmental virus contamination, the role of bovine feces in VACV transmission to rodents is unknown. The aim of this work was to evaluate the potential of cattle feces in the transmission of VACV to rodents and discuss the role of this potential viral transmission route in VACV circulation in Brazil.
Materials and Methods
To evaluate the potential of cattle feces in the transmission of VACV to rodents, we attempted to simulate this potential viral transmission route. Lactating cows were inoculated by scarification on the teats with 107 plague forming unit (PFU) VACV strain GuaraniP2 virus (GP2V) (Trindade et al. 2006, Guedes et al. 2010). Feces samples were collected daily and then examined by polymerase chain reaction (PCR) for VACV-DNA detection.
After confirming the presence of GP2V-DNA in infected cows (Fig. 1A), 2.3 g of bovine feces were diluted in 23 mL of sterile phosphate-buffered saline. This solution of feces was mixed with 42 g of wood-shavings and added to a mouse cage, containing individualized airflow. A total of five 4-week-old Balb/c mice were added to the cage and kept in direct contact with the wood shavings containing feces of cattle experimentally infected with VACV-GP2V. A control group, also with five mice, was kept in a different cage with the feces of VACV-uninfected cows. Mice of both groups were kept in their cages for 20 days without changing the wood shavings. The experiments were performed in accordance to the rules of the UFMG Ethics Committee (CETEA), at the animal facilities of our Department (UFMG).
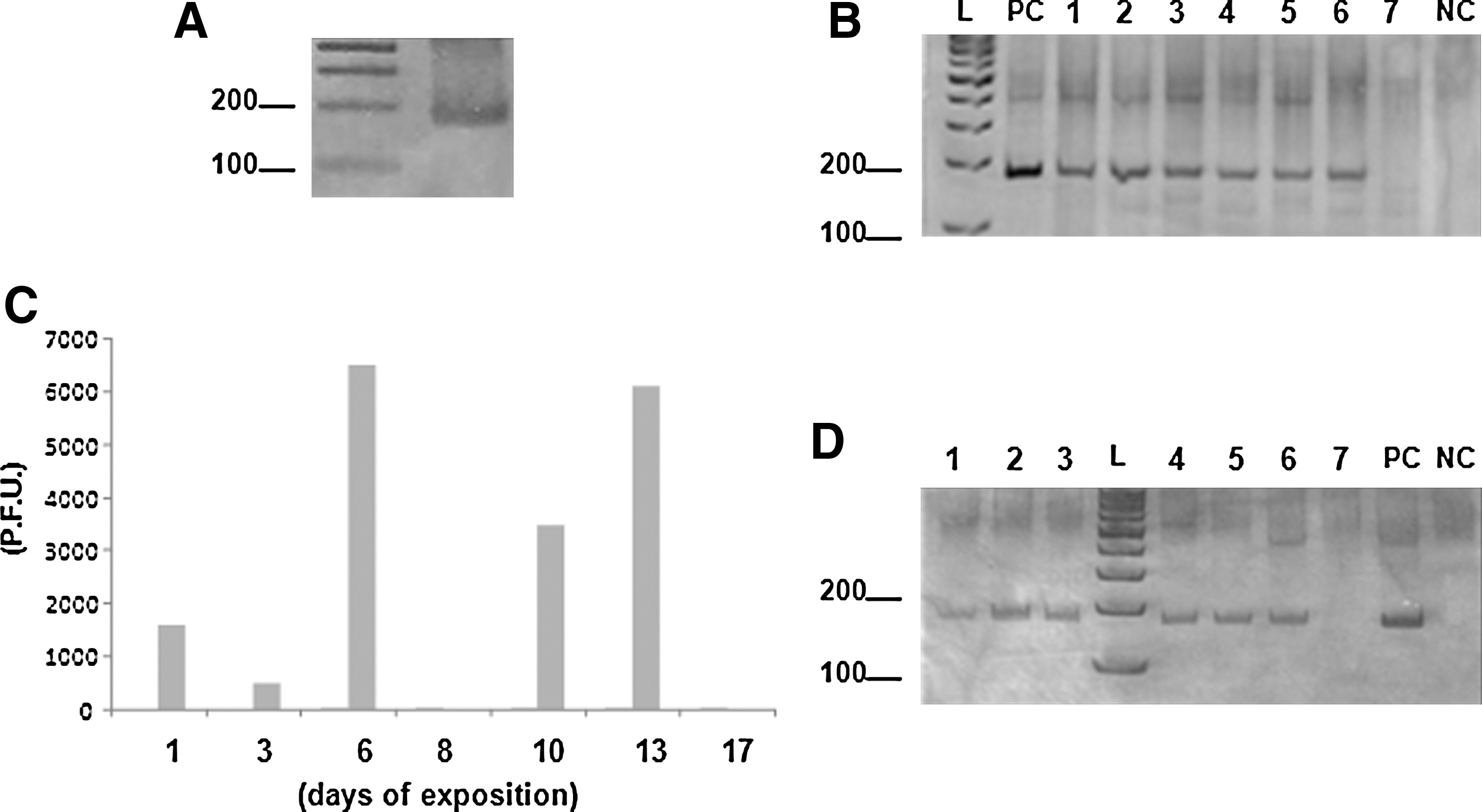
Pools of blood samples (five animals) were collected during the exposure period through a small incision at the tip of the tail. Tail bleedings were performed on alternate days, from two animals randomly chosen from each cage. In parallel with blood collection, the mice were weighed, and pools of fecal samples were collected. Feces were collected using sterile microtubes, positioned directly in the anus of the animals. These samples were then analyzed by PCR assays for VACV-vgf (Abrahão et al. 2009b) and by virus titration in BSC-40 cells (50-fold diluted). In addition, the feces and blood samples were submitted to a positive/negative (no-quantitative) real-time PCR for VACV-ha gene (Trindade et al. 2008). The animals were sacrificed after 20 days; spleen, lung, and intestine samples were collected for virus titration in BSC-40 cells. Serum samples were also collected for neutralization assays (Newman et al. 2003). All the tests were performed in duplicate.
Results
The PCR for ha and vgf genes revealed the presence of viral DNA in the blood and feces of mice exposed to feces from cattle infected with VACV-GP2V (Fig. 1B, D). VACV-specific fragments of 190 bp (vgf) were amplified from blood and fecal samples collected from the 1st to the 13th day postexposure, including a PCR positive control (Fig. 1B, D). No viral DNA was detected after day 13 postexposure. Similar results were obtained in real-time ha gene assays (data not shown). None of the fecal or blood samples collected from the control group showed amplification of virus-specific DNA fragments.
VACV infectious particles were detected in murine feces at days 1, 3, 6, 10, and 13 postexposure, at titers ranging from 5.2×102 PFU/mg (day 3) to 6.5×103 PFU/mg (day 6) (Fig. 1C). Infectious particles were not detected at day 8 and after day 13 of exposition. Spleen, lung, and intestine samples of mice exposed to feces of bovines experimentally infected with VACV were titrated. None of the organs tested showed evidence of VACV infectious particles at day 20 postexposure, although VACV DNA was detected by PCR in some intestine samples (not shown). The mice exposed to VACV-contaminated feces presented high titers of specific neutralizing antibodies, with the average of 1/80 neutralizing units per milliliter. Clinical signs were not observed in either group, likely due to the low virulence of GP2V as previously demonstrated by Ferreira and colleagues (2008a).
Discussion
In the last 11 years, our group and others have been developing epidemiological studies aiming at understanding the natural life cycle of VACV. However, the broad VACV host range makes delineation of a comprehensive and solid model difficult, because the possibilities of hosts, reservoirs, and methods of transmission seem to be countless. Despite such numerous variables, significant advances have been achieved, and some gaps are being filled. VACV transmission between humans and cows by direct contact (milking) is well known and accepted (Leite et al. 2005, Lobato et al. 2005, Silva-Fernandes et al. 2009). Transmission among cows and calves probably occurs by suckling (Leite et al. 2005, Lobato et al. 2005, Silva-Fernandes et al. 2009). Moreover, wild and peridomestic animals have been inserted into the VACV ecological puzzle, especially rodents (Abrahão et al. 2009a). How VACV can be transmitted between cattle and rodents remains a major question.
Once the viral shedding in feces of VACV experimentally infected cows was confirmed (Guedes et al. 2010), we speculated about the importance of this viral source in rodent infections. This was especially of concern, because VACV horizontal transmission by feces was previously shown in the Balb/c mouse model (Ferreira et al. 2008b). Some studies demonstrated that VACV circulates among rodents in Brazilian rural and wild areas (Lopes et al. 1965, Fonseca et al. 1998, Abrahão et al. 2009a). Rural mice and rats, which have free movement between farms and wilderness areas, usually keep direct contact with cattle feces through inhalation and ingestion of fecal material (many rodent species exhibit coprophagic behavior) (Abrahão et al. 2009a). Therefore, if the model of horizontal transmission described here could be applied to farms and wilderness, bovine feces might be considered a possible transmission source for rodent infection by VACV.
The detection of viral DNA in the blood of infected Balb/c mice without clinical signs corroborated what was observed in a previous study (Ferreira et al. 2008b). Ferreira et al. (2008b) found that mice infected with different VACV that had been previously isolated during BV outbreaks did not have clinical signs. These results reinforce the idea that rodents could play an important role in the transmission of VACV in a silent and independent way. Although there are no experimental studies related to pathogenesis of Brazilian VACV in wild rodent species, the notifications of VACV isolation from rodents with no apparent clinical symptoms in Brazilian forests (Fonseca et al. 1998, Abrahão et al. 2009a) reinforce this theory. Despite the lack of information regarding VACV pathogenesis in wild rodents, viremia could also be a common phenomenon in VACV infected rodents, once the VACV BeAn58058 virus was isolated from blood of the Amazonian wild Oryzomys sp. rodent (Fonseca et al. 1998). The absence of infectious particles detection in organs of mice exposed to GP2V-contaminated bovine feces are compatible with previous studies, as GP2V is a low virulent VACV strain (Ferreira et al. 2008a). The unique organ that VACV GP2V infectious particles were detected in intranasally infected and sentinel mice is the intestine, as described by Ferreira et al. (2008b); however, in that study, the titration assays were performed at day 10 post-infection in contrast with the present study, in which the organs were tittered at day 20 of exposition. New related aspects of VACV transmission still need to be explored, including pathogenesis studies in wild rodents. Such studies could help explain how the absence of the verification for VACV propagation in experimentally infected animals could influence the VACV circulation and transmission among rodents. Research on VACV circulation is a wide and open area, and it is related to important aspects of public health. The social, economical, and medical impacts associated with BV outbreaks in Brazil encourage studies on VACV host range, pathogenesis, and epidemiology. Each piece of the VACV puzzle has great importance in the planning for control and prevention of this emerging zoonotic disease. Bovine feces could be an important piece in this puzzle, as a possible transmission source for rodent infection by VACV.
Footnotes
Acknowledgments
The authors thank MSc. João R. dos Santos, Angela S. Lopes, Ilda M.V. Gama, Andreza de Carvalho, and colleagues from Laboratório de Vírus (ICB-UFMG). Financial support was provided by CNPq, CAPES, FAPEMIG, and MAPA.
Disclosure Statement
No competing financial interests exist.