
Abstract
The decline in the number of northern fur seal (NFS; Callorhinus ursinus) pups on St. Paul Island, Alaska, has led to multidisciplinary research, including investigation into issues of reproductive health and success. Given the recent identification of Coxiella burnetii in the placenta of two other marine mammal species, NFS placentas were collected from Reef rookery on St. Paul Island, Alaska, during the 2010 pupping season, examined histologically, and tested for C. burnetii using polymerase chain reaction (PCR). Of 146 placentas examined, gram-negative intratrophoblastic bacteria that were positive for C. burnetii on immunohistochemistry were observed in 5 (3%) placentas. Placental infection was usually devoid of associated inflammation or significant ancillary pathology. One hundred nine (75%) of the placentas were positive for C. burnetii on PCR. C. burnetii is globally distributed and persists for long periods in the environment, providing ample opportunity for exposure of many species. The significance of this finding for the declining fur seal population, potential human exposure and infection, and impact on other sympatric marine mammal or terrestrial species is unclear; further investigation into the epidemiology of Coxiella in the marine ecosystem is warranted.
Introduction
Coxiella burnetii is a gram-negative bacterium with a global distribution. The organism has an extremely broad host range, including humans, wild and domestic mammals, birds, and arthropod tick vectors (McQuiston et al. 2002). C. burnetii has recently been identified in the placenta of a single Pacific harbor seal in California (Lapointe et al. 1999) and a Steller sea lion in Washington State (Kersh et al. 2010a). However, the prevalence and significance of this bacterium in the marine ecosystem is largely unknown. Mild placentitis was identified in 21 percent of premature, stillborn, or neonatal mortalities in NFS between 1986 and 2006 when histopathology was conducted (Spraker and Lander 2010), suggesting the possibility of placental infectious disease such as C. burnetii infection in this species. The objective of this study was to determine the prevalence of C. burnetii infection in NFS placentas.
Materials and Methods
From July 10 to August 10, 2010, fresh placentas were collected twice daily from Reef rookery on St. Paul Island, Alaska. Placentas were picked up using 10 m bamboo poles with hooks from catwalks above the rookery. Placentas were evaluated grossly for pathologic lesions and representative sections were collected fresh and frozen at −20°C as well as fixed in 10% neutral buffered formalin. Two to four representative areas of fixed placenta were paraffin-embedded, and tissue blocks were sectioned at 5 μm and stained with hematoxylin and eosin for histopathologic evaluation.
On tissues in which intratrophoblastic organisms were observed, immunohistochemical staining for C. burnetii was performed using standard manual techniques on a vertical slide staining system (Sequenza; Thermo Electron Co., Pittsburgh, PA). Briefly, 5-μm sections were mounted on positively charged slides, deparaffinized, and then rehydrated with descending alcohol concentrations to buffer. Antigen retrieval was performed using an ethylenediaminetetraacetic acid-based solution, pH 9.0 (S2367; Dako Cytomation, Carpinteria, CA) and incubating at 125°C for 1 min. The primary antibody was C. burnetii–positive human control serum (CC007; Panbio Diagnostics, Columbia, MD), incubated at a 1:50 dilution overnight at 4°C. The secondary antibody, biotinylated goat anti-human (ab6857; Abcam; Cambridge, MA), was applied for 1 hour at a 1:1000 dilution, and followed with a streptavidin–horseradish peroxidase conjugate (SA-5004; Vector Laboratories, Burlingame, CA) for 20 min at 1:1500 dilution. Immunoreactive complexes were detected using 3-amino-9-ethylcarbazole substrate (SK-4200; Vector Laboratories; Burlingame, CA) followed by counterstaining with Mayer's hematoxylin. C.burnetii-positive Steller sea lion placenta (Kersh et al. 2010a) was used for a positive tissue control slide, and the primary antibody was replaced with antibody dilutant for a negative antibody control slide.
For polymerase chain reaction (PCR), DNA was purified from approximately 330 μg of frozen placenta tissue using the Qiagen QIAamp DNA mini kit tissue protocol (Qiagen, Valencia, CA). The PCR assays for IS1111a and com1 were quantitative TaqMan assays that have been reported previously (Kersh et al. 2010a). Standard curves using 10-fold dilutions of Nine Mile Phase 1 C. burnetii DNA were run for quantitation. Oligonucleotides were synthesized by the biotechnology core facility at the Centers for Disease Control, Atlanta, GA.
Results
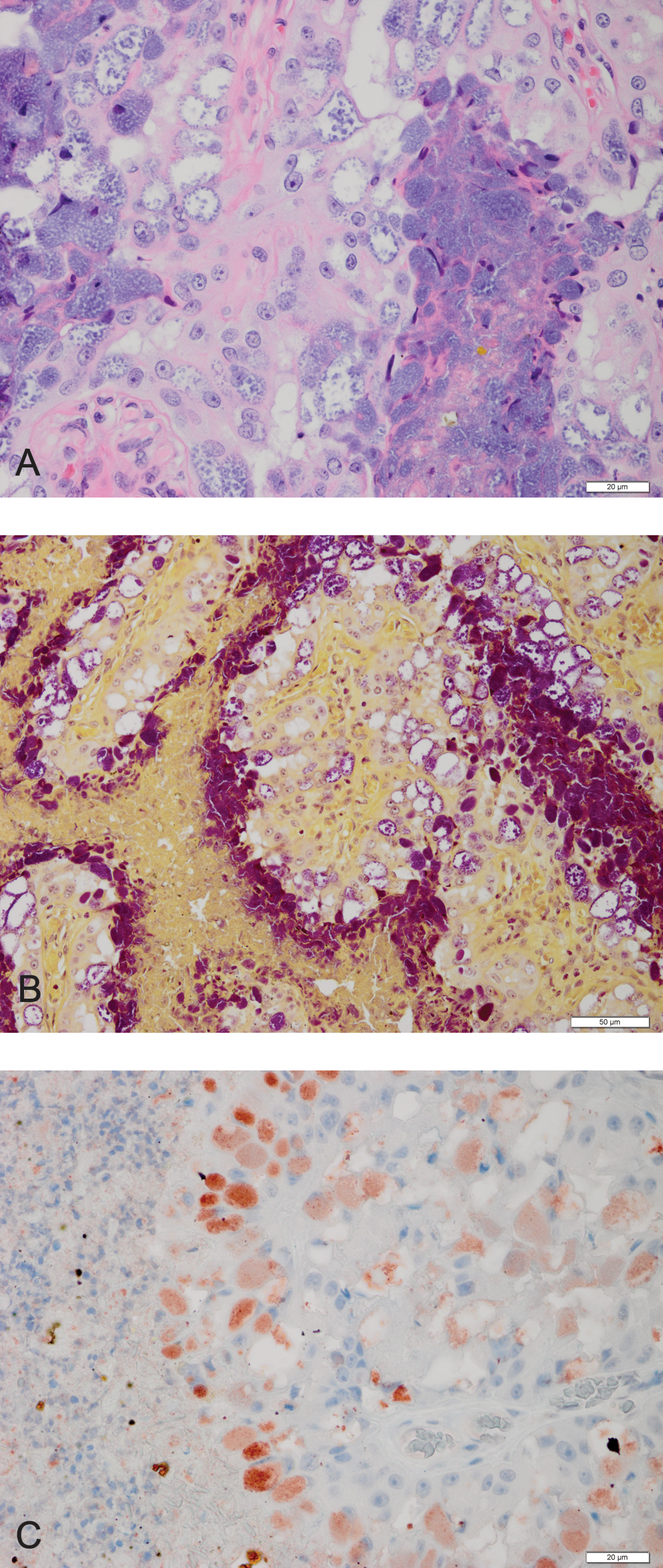
One hundred forty-six placentas were retrieved and no gross lesions were identified. All placentas had an obvious hemophagous zone at the margin of the placenta with smaller, similar-appearing hematomas distributed randomly throughout the tissue. In all placentas, there were mild to moderate amounts of necrotic cellular debris associated most commonly with superficial chorionic villi but also present within crypts. In some cases only cellular debris was identifiable, but elsewhere cellular apoptosis could be identified. Placental mineralization was present in 60 (41%) of the tissues. Five placentas (3%) had regionally extensive areas of trophoblastic, intracytoplasmic bacteria (Fig. 1A). Organisms were gram negative (Fig. 1B) and immunohistochemically positive for C. burnetii (Fig. 1C). Infection of trophoblasts was patchy and not present in all sections examined from the five IHC positive tissues. Areas of infection were usually devoid of associated inflammation, however fibrinous, mononuclear arteritis of a large vessel was identified in some sections of a single placenta. Tissue degeneration and necrosis was not increased in areas of infection relative to elsewhere within the tissue, with the exception of sections in which arteritis was observed.
To confirm the identity of the organisms, DNA was purified from the same set of 146 placentas, and quantitative PCR for the C. burnetii–specific genes IS1111a and com1 was performed. All 146 placentas were negative for IS1111a, but 109/146 (74.7%) were positive for com1. The genome equivalents per gram of tissue for the 109 com1-positive placenta samples are shown in Figure 2. For the five placentas that had infection confirmed by IHC, the densities of organism (per gram) detected by PCR were 3×105, 1×106, 1.2×106, 3.5×106, and 8.4×108; a total of 25 (17%) samples had organism counts ≥3×105/g.

Number of polymerase chain reaction-positive placentas with different amounts of C. burnetii DNA among the 109 com1-positive placenta samples.
Discussion
Results of this study suggest that NFS placentas are frequently infected with C. burnetii. The reason for the discordant test results between PCR and IHC is not completely understood. Organisms observed microscopically were not uniformly distributed within or between sections taken from each IHC-positive placenta, and the focal nature of the infections could account for many infected tissues being missed on histology. Additionally, the high molecular prevalence may have been influenced by environmental contamination of some placentas. In addition to birth products, C. burnetii can be shed in milk, urine, and feces. The high density of animals present on the rookery opens the possibility that PCR-positive results for some placentas could be due to contamination with C. burnetii derived from infected animals in proximity. For the 40 PCR-positive placentas that had <104 genome equivalents per gram, environmental contamination is a possibility. The IHC and PCR results were compared, and it is assumed that placentas with >105 genome equivalents per gram were likely to have a genuine infection, but a better understanding of diagnostic test performance in this species is a critical part of investigating the epidemiology of Coxiella infection in the marine ecosystem. Although most strains of C. burnetii have an IS1111a gene, poor amplification of IS1111a has previously been observed in a C. burnetii strain infecting a Steller sea lion placenta (Kersh et al. 2010a). This suggests that the strains infecting the NFSs may be related to strains infecting other marine mammals and that an altered form of IS1111a may be a feature of these strains.
The histology of seal placentas has been well described in previous reports (Rowlands 1966) and histologic anatomy in this set of NFSs is consistent with those descriptions. As with most carnivores, placentation is best described as circular, zonary, labyrinthine, non- or mildly-deciduate and endotheliochorial. The hemophagus organ is characterized by phagocytosis of red blood cells as part of normal uptake of iron for the developing fetus (Rowlands 1966). The most common histologic feature in all of the NFS placentas was evidence of apoptosis and necrosis—both features routinely identified in normal placentas as part of physiologic detachment and separation during parturition. Apoptosis or programmed cell death is common in normal, physiologic cell turnover; in human placentas, apoptosis is more common in the third trimester, presumably as the fetus nears term and normal parturition (Smith et al. 1997). Increased apoptosis has been described in disease states; however, there was no other histologic evidence to suggest that the process was pathologic in the study population. Placental mineralization, identified in almost half of the examined placentas, is also considered a common and normal finding in animal placentas (Maxie 2007).
Histologically, placental infection with C. burnetii was consistent with that described previously in seal and sea lion placentas (Lapointe et al. 1999, Kersh et al. 2010a). Given the global distribution of the organism and its persistence in the terrestrial environment, the two diseased marine mammals in previous reports may have been exposed to the bacteria in a number of locations and represent sporadic cases of clinical disease. The high molecular prevalence reported here suggests that NFS may be capable of maintaining the infection within the population. Of interest was the notable lack of significant pathology in most of the infected tissues suggesting either a host adaptation to the organism or simply that the lesion distribution and severity is variable both between and within placentas. Although individual animal identification was not possible on the rookery, all NFS placentas were presumed to be from healthy births as they were not collected near a dead pup. As such the significance of infection is difficult to discern. In terrestrial livestock, placental infection with C. burnetii culminating in abortion is commonly associated with severe placentitis and often vasculitis (Bildfell et al. 2000, Sánchez et al. 2006); however, within endemic small ruminant herds, the degree of inflammation can be variable. It is possible that more severe infections in NFS that lead to severe placentitis or placental detachment could result in abortion at sea, and would not be observable in samples collected at the rookery. While placentitis has been described previously in NFS placentas, no infectious etiology was identified in those cases and histopathology was not consistently done on all cases, making interpretation of historical frequency data challenging (Spraker and Lander 2010).
The role of marine mammals in the epidemiology of Coxiella is unclear. In the United States, sheep and goats are thought to represent the greatest risk to humans although evidence of exposure and infection is present in a wide range of domestic and wild species (McQuiston and Childs 2002). In ruminants, the bacteria can infect the uterus asymptomatically and be shed in abundance at parturition, contaminating the environment (Maurin and Raoult 1999) where it is extremely persistent (Arricau-Bouvery and Rodolakis 2005) and providing opportunities for exposure and infection including through aerosols (Astobiza et al. 2010). In the United States the organism has been isolated from the environment in both agricultural settings as well as areas with predominantly human activity (Kersh et al. 2010b). A single organism can result in severe human disease, and the organism has therefore been identified as a potential bioterrorism agent (Anderson et al. 2006). In Greenland, seals were implicated as a possible source of exposure for a human case of Q fever (Koch et al. 2010); however, this was based on activities of the patient without knowledge of the infection status of regional seals. Given that subsistence harvesting is conducted within this population of Alaskan NFS, further research into the pathogen, prevalence, and shedding of the organism as a source of human exposure may be warranted.
The relationship between C. burnetii infection and the declining pup production that has occurred on St. Paul Island since 1998 is, at this point, unknown. Pregnancy rates for premigratory seals have been shown to be high (≥85%), unlikely to have changed since before the current decline (Testa et al. 2010) and there has been no documented increase in the presence of aborted or premature fetuses on the rookery where the placentas examined for this study were collected. NFSs have not previously been tested for C. burnetii on St. Paul Island or any other known rookeries, thus exposing the need for additional studies to determine if this organism represents a current pathogenic threat to the population or if it is a commensal organism that has been present long before recent population declines.
Footnotes
Acknowledgments
The authors would like to thank Juan Leon Guerrero, Bobette Dickerson, Mike Williams, and Rolf Ream for their help collecting placental tissue and insightful discussion on fur seal biology and disease. All tissue samples were collected under authority of U.S. Marine Mammal Permit No. 782-1708 issued to the National Marine Mammal Lab, Seattle.
Disclosure Statement
No conflicts of interest or competing financial interests exist on the part of any authors.