
Abstract
Yellow fever (YF) is a viral hemorrhagic fever endemic to tropical regions of South America and Africa. From 2007 to 2009 an important epidemic/epizootic of YF was detected in different populations of howler monkeys (Alouatta species) in Misiones, a northeastern Argentinian province. Yellow fever virus (YFV) infection was researched and documented by laboratory tests in humans and in dead Alouatta carayá. The objective of that research was to investigate the circulation of YFV in mosquitoes, which could be implicated in the sylvatic transmission of YF in Argentina. The above-mentioned mosquitoes were captured in the same geographical region where the epizootic took place. A YFV strain was isolated in cell culture from pools of Sabethes albiprivus. This study is not only the first isolation of YFV from mosquitoes in Argentina, but it is also the first YFV isolation reported in the species Sabethes albiprivus, suggesting that this species might be playing a key role in sylvatic YF in Argentina.
Introduction
It is considered that this virus originated in Africa and was introduced into the New World in slave ships bearing active cases of YF by infected Aedes aegypti mosquitoes (Bryant et al. 2007). The enzootic transmission cycle involves two components: the mosquitoes and the nonhuman primates. In Africa the vector involved in this enzootic cycle is a mosquito from the genus Aedes, whereas in South America the vectors associated with the transmission cycle are mosquitoes from the genera Haemagogus and Sabethes (Monath 1999; Vasconcelos 2003; Camargo-Neves et al. 2005).
Currently, nucleotide sequencing studies of structural gene regions and 3′NCR delineated seven genotypes of YFV worldwide: five genotypes in Africa, and two in South America. In this context, the Brazilian and Peruvian YFVs represent the two major South American YFV genotypes I and II, respectively (Bryant et al. 2003b). Specifically in Argentina, the last YFV outbreak was detected in Corrientes and Misiones provinces in 1966, in which a total of 53 human cases were notified: 41 in Misiones province and 12 in Corrientes province. However, only five cases were confirmed by means of virological and/or histopathological studies. At that time, the mosquitoes collected in the field yielded negative results (Sabattini et al. 1998). More recently, a YFV outbreak was seen in the same provinces from November 2007 to February 2009 (Holzmann et al. 2010), involving the illness and death of humans and howler monkey populations. In this outbreak, YFV was confirmed by laboratory tests in 9 human cases and 16 dead howler monkeys (unpublished data). Nevertheless, the role of mosquitoes as potential vectors in the transmission cycle of this epizootic is still unknown. Therefore, the main goal of this study was to investigate mosquito species that could play a role in the sylvatic transmission of YFV in Argentina.
Materials and Methods
Study areas, collection methods, and identification of exemplars
Mosquito captures were carried out in Misiones province, Argentina (Fig. 1) from January 7 to January 14, 2009. The sampling localities were selected based on the places where YFV was confirmed in dead howler monkeys. One of the sampling sites was Garupá, which is a rural area 16 km away from Posadas city (27° 29’65” S/55° 52’52” W), with a population of around 70,000 inhabitants. The second site was Tres Capones (28° 01’00” S/55° 37’22” W), a rural area 60 km away from Posadas city with 1234 inhabitants. Both sites have similar climatic conditions, characterized by a mean annual temperature of 21.5°C (range 15–35°C), high humidity (74%), and annual rainfall averages 2000 mm. Forests in this region are classified as deciduous stationary forest because of the low temperatures during the winter season. The landscape is dominated by plantations of yerba mate (Ilex paraguariensis) and ranching, which demonstrates the extensive human influence on the environment. The sites are characterized by a mosaic of vegetation surrounded by areas dedicated to cattle breeding and forest in galleries along rivers. Monkey populations live in the gallery forests where they can find water and food.

Location of the capture area.
Capture of adult mosquitoes was carried out with CDC traps. The traps operated for 24 continuous hours in each site, in periods from 8
In addition, manual capture was performed at ground level and at the forest canopy level at an elevation of approximately 10–15 m, depending on the local trees and vegetation, by using entomologic net and bottle-type manual vacuums. The collections took place from 9
Insects collected in the field were placed in labeled tubes, frozen in liquid nitrogen, and shipped to the Instituto Nacional de Enfermedades Virales Humanas (INEVH) laboratory. Upon arrival at the laboratory, the specimens were sorted under a stereoscopic microscope on a chilled table by species, capture method, location, and date of collection. Due to the damage suffered during the collection and shipping, many specimens were only identified to the genus level, and when possible, they were also identified to the level of species. Morphological identification of all species was done by means of the key from Darsie (1985). In addition, a molecular characterization of the specimens was performed using the mitochondrial gene cytochrome I (COI) as genetic marker. DNA was extracted from two legs of each individual using a standard phenol-chloroform method. A 445-bp segment was amplified by standard PCR procedures, using the universal insect primer CI-J-1718 (forward). and a reverse primer designed as the complementary sequence of the CI-J-2183 (Simon et al. 1994). Sequences were obtained using ABI PRISM 3100 DNA Analyzer equipment. The COI sequences of virus-positive mosquitoes were deposited in the GenBank database under accession numbers GU992947, GU992948, GU992949, GU992950, and GU992951.
Virus isolation
Pools of mosquitoes were triturated in sterile grinders containing 1 mL of phosphate-buffered saline solution with 0.75% bovine albumin, penicillin (100 units/mL), and streptomycin (100 μg/mL). The homogenate was centrifuged at 14,000 rpm for 10 min. A 0.2-mL aliquot of each original suspension of mosquitoes from the genus Sabethes or Haemagogus was inoculated into a mammalian cell line derived from monkey Vero C/76 cells, and in a cell line derived from Aedes albopictus C6/36 in order to isolate the YFV. Cell cultures were observed daily for cythopatic effect for 14 days. Virus isolates were identified by inmunofluorescence using specific monoclonal antibodies against YFV (Mab FA 2D12).
The minimum infection rate (MIR) of the mosquitoes was calculated as the ratio of the total number of YFV-positive pools to the total number of mosquitoes processed of that species×100 (Walter et al. 1980; Katholi and Unnasch 2006).
Reverse-transcription PCR and sequencing
RNA was extracted using TRI Reagent™ (Sigma-Aldrich, St. Louis, MO) from original suspensions of mosquitoes and from cell culture supernatants of the original isolations. After that, a reverse-transcription polymerase chain reaction (reverse-transcription PCR) was performed to amplify YFV-specific nucleic acid by using primers previously described in Sánchez-Seco and associates (2006). The amplicons of the expected size (505 bp) were cut from the agarose gel, purified by a QIAquick kit (Qiagen, Valencia, CA) according to the manufacturer's protocol, and finally sequenced directly from both strands for verification using ABI PRISM 3100 DNA Analyzer equipment.
In order to analyze the YFV genotype, sequence data were obtained from fragments of two genomic regions, one spanning the terminal portion of NS5 and the proximal region of the 3′NCR using primers EMF (5′TGGATGACKGARGAYAT) and VD8 (5′GGGTCTCCTCTAACCTCTAG; Vasconcelos et al. 2004), and a second fragment from the genomic region comprising the premembrane (prM) and envelope (E) glycoprotein genes, using the genomic-sense primer (5′CTGTCCCAATCTCAGTCC), and the genomic-complementary primer (5′AATGCTTCCTTTCCCAAAT; Bryant et al. 2003a). The amplicons of the expected size (595 bp and 670 bp, respectively) were cut from the agarose gel, purified using a QIAquick kit, according to the manufacturer's protocol, and sequenced directly from both strands of each reverse-transcription PCR product for verification using ABI PRISM 3100 DNA Analyzer equipment.
Phylogenetic analysis
A fragment of 670 bp corresponding to the prM/E region and other fragment of 595 bp corresponding to the NS5/3′NC region were sequenced and deposited in the GenBank nucleotide sequence under accession numbers HQ123568; HQ123569; HQ123570; HQ123571 (preM/E); HQ123572; HQ123573; HQ123574; HQ123575 (NS5/3′NCR).
The sources, geographic origins, and GenBank accession numbers of the prM/E junction and the NS5/3′NCR sequences of YFV used in the phylogenetic analysis are listed in Table 1.
AP, Amapa; BA, Bahia; GO, Goias; MG, Minas Gerais; MS, Mato Grosso do Sul; PA, Pará; RO, Rondônia; RR, Roraima; SP, Sao Paulo; TO, Tocantins; ND, no data.
Sequences were edited and aligned with the BioEdit program and the Clustal Wallis method (available from
The phylogeny of the sequences was constructed using the Bayesian analysis, Neighbor Joining, and Parsimony methods. Bayesian analysis was performed using MR. BAYES 3.1.2 (Ronquist and Huelsenbeck 2003). The best model suggested by jModeltest 01.1 was TIM2 (Posada 2008). Because of limited availability of models in MR. BAYES, the selected parameters used were: number of substitutions (nst) 2, and rates inv gamma. The Markov Chain Monte Carlo was 5,000,000 generations long, sampling every 100 generations, for a total of 50,001 samples. Of these, the first 2500 were discarded as burn-in to assure reaching a plateau. Maximum parsimony analysis was performed using the TNT v1.0 program (Goloboff et al. 2000), and Jackknife support values were obtained using 1000 replicates. Neighbor Joining trees were constructed with MEGA 5 Software (Tamura et al. 2011). The nucleotide distance was calculated by using the Kimura-2+G method and bootstrap analyses with 1000 pseudoreplicates.
In all cases, to enable better rooting of the phylogenetic tree, the strains Car77, Asibi, and a strain of YFV from Uganda were added as outgroups.
Results
During the study period, a total of 495 mosquitoes were captured: 395 at Garupá and 100 at Tres Capones. They were sorted into 128 pools (98 pools from Garupá mosquitoes and 30 pools from Tres Capones mosquitoes).
The sample obtained in Garupá was larger than the one obtained in Tres Capones for both capture methods (Table 2). However, at both sampling sites, the great majority of mosquitoes were caught with CDC traps. In addition, the number of mosquitoes captured on the forest canopy level was lower than on the ground level. It is important to mention that the sampling effort was mainly focused on the ground.
Number mosquitoes captured (number of pools).
Pools tested for virus isolation attempt.
Culex mosquitoes were the most abundant genus, followed by Sabethes. The predominant species among the Sabethes genus were Sa. albiprivus and S. chloropterus. Manual capture was more successful than CDC traps for trapping Sabethes mosquitoes, including Sa. albiprivus. Specimens of genera Aedes, Shannoniana, and Wyeomyia were less representative.
All of the 128 pools were tested for flavivirus by reverse-transcription PCR. In addition, the 73 pools belonging to genera Haemagogus and Sabethes were also tested for virus isolation (Table 2).
The 50 captured specimens of Sa. albiprivus were sorted into 20 pools, and one of them was positive for YFV with the two techniques used. The YFV strain was isolated in Vero C/76 and C6/36 cell cultures. Both lines of cells showed cytopathic effect at day 6 after inoculation. This positive pool included seven individuals collected in Garupá. The calculated MIR was 2%. None of the remaining pools showed any positive results.
The Bayesian analysis of the nucleotide sequence of a 670-bp fragment of a genomic region prM/E (Fig. 2), and a 595-bp fragment of the NS5/3′NCR (data not shown), demonstrated that the YFV strain isolated from Sa. albiprivus is positioned together with other YFV strains isolated from Argentina and Brazil in 2008.
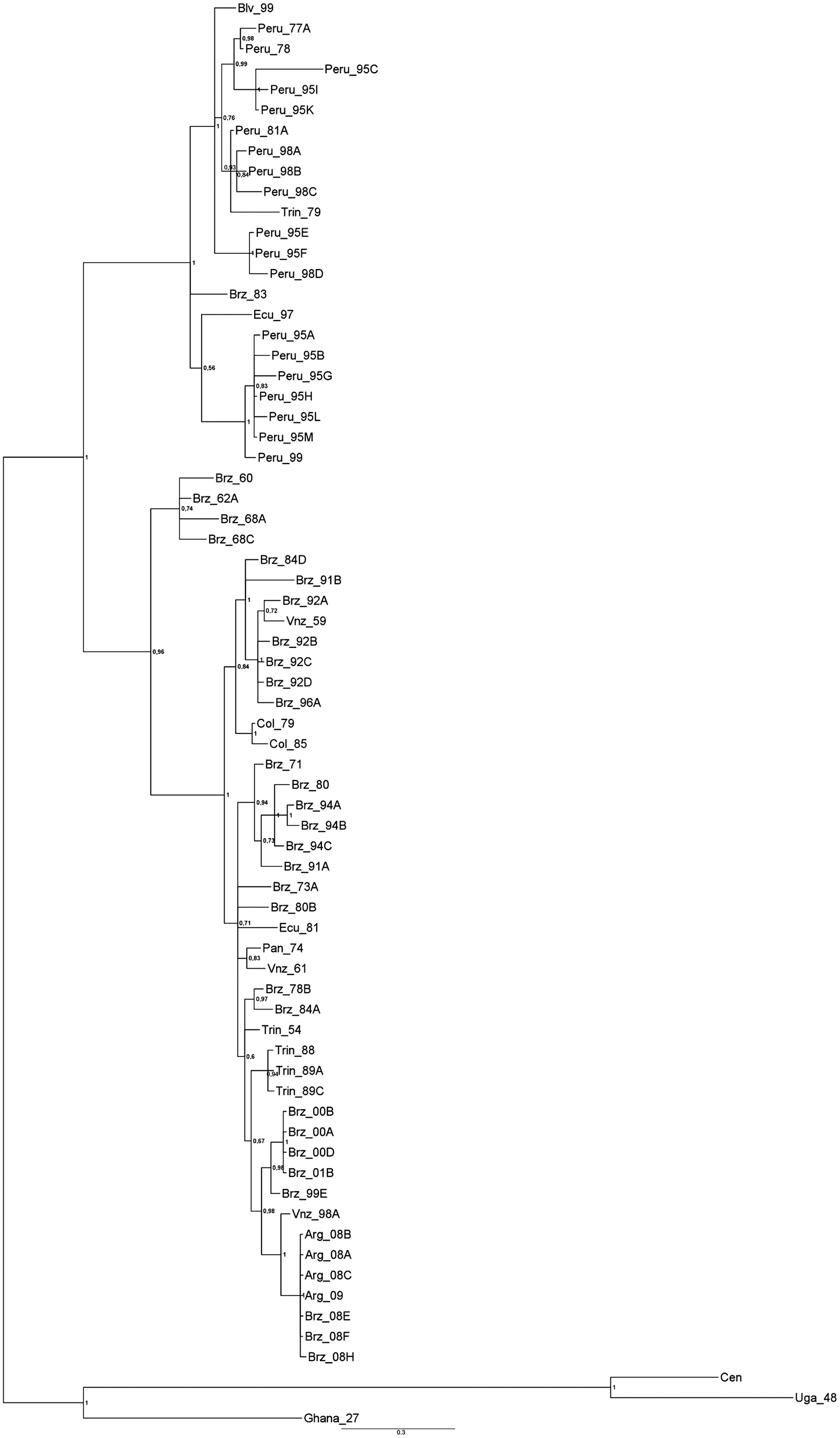
Bayesian analysis based on a 670-bp fragment from the yellow fever virus (YFV) genome structural region prM/E. This analysis used the TIM2 model, as suggested by jModeltest. Numbers above the nodes represent posterior probabilities of those branches. The sequences of YFV from Uganda, Ghana, and the Central African Republic were used as outgroup (Arg, Argentina; Brz, Brazil; Trin, Trinidad; Vnz, Venezuela; Col, Colombia; Pan, Panamá; Ecu, Ecuador, Blv, Bolivia; Cen, Central African Republic; Uga, Uganda).
Discussion
Between 2007 and 2009, sylvatic YF outbreaks were documented in northeastern Argentina. From December 2008 to February 2009, several deaths among howler monkeys (A. carayá) were reported in southern Misiones province. Naturally YFV acquired infections in the neotropical forest may result in disease and death in monkey population; indeed, the finding of dead howler monkeys (Alouatta sp.) has signaled the occurrence of a YF epizootic (Monath 1989).
Previous studies reported the isolation of YFV from human cases of a YF outbreak that occurred in 1966 in Corrientes and Misiones provinces, Argentina (Barrera Oro et al. 1966). Since then, no human or monkey cases have been reported, and thus our knowledge of sylvan vectors of YF in Argentina is scarce.
Haemagogus and Sabethes mosquitoes are distinctly diurnal biters, with peak activity seen during the afternoon, and previous studies about host preference clearly show that these genera are strongly attracted to humans (Monath 1989; Castro Gomez et al. 2010). Therefore, this behavior may explain the fact that manual capture was more successful than CDC traps for trapping Sabethes mosquitoes, including Sa. albiprivus.
All mosquitoes captured were studied for flavivirus detection using reverse-transcription PCR; however, Sabethes and Haemagogus genus mosquitoes were selectively tested for YFV because these genera have been associated with transmission of YFV in sylvatic areas in South America (Monath 1999; Vasconcelos 2003; Camargo-Neves et al. 2005). In 2001 and 2008 strains of YFV were isolated from Haemagogus leucocelaenus, and from Aedes serratus in Rio Grande do Sul State, Brazil (Vasconcelos et al. 2003; Cardoso et al. 2010). Contrary to this finding, our study did not find any Ae. serratus in the sampling sites, and the population of Hg. leucocelaenus was very small. Sabethes was the most abundant genus after Culex, and Sa. albiprivus was the most representative species of the Sabethes mosquitoes. In fact, the geographic distribution of Sa. albiprivus includes Bolivia, Brazil, Argentina, Colombia, French Guiana, Paraguay, and Suriname.
Vectorial capacity of mosquitoes involved in YF transmission is profoundly affected by rainfall and temperature, which in turn follow definite seasonal patterns depending on latitude. The 2008–2009 summer was dry in the area of the outbreak. Mosquitoes from Sabethes genus had been implicated in the transmission of YFV in areas subjected to a prolonged dry season during which Haemagogus activity subsides; it is worth mentioning that mosquitoes from this genus are drought-resistant and long-lived (Monath 1989).
In the Americas, Haemagogus janthinomys, Hg. leucocelaenus, Hg. lucifer, and Sabethes chloropterus are captured preferentially in tropical rain forest habitat. In undisturbed forests their activity is often highly restricted to the canopy. However, where the forest is disturbed or replaced by lower trees and more open conditions (as in grassland-gallery forest areas), the vertical distribution of these species may change, increasing their activity at ground level. Moreover, deciduous forests associated with a pronounced dry season, such as the sampling localities, show less vertical stratification of all vector species, and the activity at ground level increases (Monath 1989).This may be one of the reasons why so few individuals were captured at the canopy level.
Different studies about population dynamics done in Rio Grande do Sul Brazil show small populations of mosquitoes of the genera Haemagogus and Sabethes, probably because of environmental degradation (Paterno and Marcondes 2004; Castro Gomez et al. 2009, 2010; Reis et al. 2010).
From 2007–2009 epidemic and epizootic outbreaks of YF occurred in Argentina, and several YFV strains were isolated from humans and howler monkeys (unpublished data). The phylogenetic analyses were carried out by sequence data based on the nucleotide sequences of the prM/E region, and NS5/3′NCR (data not shown) from 70 YFV strains. They were analyzed with the use of algorithms for parsimony method, for distance method (data not shown), and for Bayesian analysis (Fig. 2). The phylogenetic trees generated by these analyses had the same topology.
The phylogenetic analysis showed that the strain of YFV isolated from Sa. albiprivus is positioned together with other Argentinean YFV strains isolated from humans and monkeys, forming a well-supported clade (Fig. 2). Remarkably, in the same period, several YF cases in humans and monkeys were also diagnosed in Brazil and Paraguay (Secretaria de Saúde do Governo do Estado do Säo Paulo, Brasil 2009; Pan American Health Organization 2008). Previous studies revealed that Brazilian and Peruvian YFV strains are significantly divergent virus lineages that can be differentiated on the basis of molecular markers in both the structural proteins and the 3′NCR. (Bryant et al. 2003b). Our results reveal that the YFV strain isolated from Sa. albiprivus from Argentina is placed with other strains from the last YF outbreak that occurred in Brazil in 2008, within the genotype I clade (Fig. 2). This genotype was first reported in Brazil, suggesting that the outbreak in Argentina may be a spillover from Brazil.
In summary, the isolation of a YFV strain from Sa. albiprivus from Argentina reported in this study is the first case of YFV isolation from mosquitoes in this country. Furthermore, it is the first time that a YFV strain has been isolated from Sa. albiprivus. In general, the entomologic vigilance is principally focused on mosquitoes from genus Haemagogus; our results indicate the necessity of studying the population of species from genus Sabethes, including Sa. albiprivus, during YF outbreaks. These findings indicate that Sa. albiprivus is a putative vector of sylvatic YF, at least in Argentina. It is possible that Sa. albiprivus plays an important role in the maintenance of YFV, although further studies are necessary to determine its effectiveness as a YFV vector in the study area.
Footnotes
Acknowledgments
The authors wish to thank the anonymous reviewer for critical suggestions and advice that helped to improve previous versions of this article. They also gratefully acknowledge the personal support of O'Duyer German, Paura Jose, and Polidoro Cesar, and all the INEVH staff, and particularly to Luppo Victoria for technical assistance, and Bonano Diego for technical support. They also thank the Ministerio de Salud de la Provincia de Misiones and Dr. Julio Estevez, the Department of Environmental Health of Posadas City, and Mr. Albarenga Luis.
Author Disclosure Statement
No competing financial interests exist.