
Abstract
We applied a process-driven model to evaluate the impact of climate scenarios for the years 2020, 2050, and 2080 on the life cycle of Hyalomma marginatum ticks in the western Palearctic. The net growth rate of the tick populations increased in every scenario tested compared to the current climate baseline. These results support the expectations of increased tick survival and increased population turnover in future climate scenarios. We included a basic evaluation of host movement based on rules connected to altitude, slope, size of the near patches, and inter-patch distances in the real landscape over the target area. Data on landscape were obtained from medium-resolution MODIS satellite imagery, which allowed us to test the potential spread of the populations. Such a model of host dispersal linked to the process-driven life cycle model demonstrated that eastern (Turkey, Russia, and Balkans) populations of H. marginatum currently are well separated and have little mixing with western (Italy, Spain, and northern Africa) populations. The northern limit is marked by the cold areas in the Balkans, Alps, and Pyrenees. Under the warmer conditions predicted by the climate scenarios, the exchange of ticks throughout new areas, previously free of the vector, is expected to increase, mainly in the Balkans and southern Russia, over the limit of the mountain ranges. Therefore, the northern limit of the tick range would increase. Additional studies are necessary to understand the implications of host changes in range and abundance for H. marginatum and Crimean-Congo hemorrhagic fever virus.
Introduction
Crimean-Congo hemorrhagic fever (CCHF) is a viral zoonosis transmitted mainly by tick bites, which is of concern in many regions of the world, including Africa, the Middle East, southern and eastern Europe, and western Asia. The virus is transmitted to reservoir mammals and humans through the bite of hard ticks, mainly of the genus Hyalomma (Hoogstraal 1979). Humans may also become infected through direct contact with the blood or tissues of infected humans or livestock (Hoogstraal 1979). Since the first outbreak of CCHF was described in Europe in 1945, several subsequent outbreaks have been reported worldwide in both newly discovered foci and foci at which the virus was known to be present (Ergonul 2006). Recent reports of an increased incidence of CCHF stimulated speculation about the presumed effects of climate on the historical geographic range of H. marginatum ticks in the Palearctic region (Karti et al. 2004; Maltezou and Papa 2010), and the probable spread of the pathogen. The tick is presumed to be the most prominent vector of the virus to humans in a large region extending from the Balkans in Europe to Pakistan and Afghanistan in the Middle East (Ergonul and Whitehouse 2007). In an expert consultation organized by the European Center for Disease Control in 2008, a short-term priority was recognized to be “endemic regions in countries with CCHF in southeastern Europe should be further mapped on national and international levels, and the degree of CCHF risk in all countries should be estimated.”
Ticks can disperse large distances only while on their hosts (Randolph 1998). Therefore, the capacity for a population to spread depends on the availability and invading abilities of the potential hosts, in combination with other factors that deeply affect the behavior of the host, such as habitat fragmentation and physical barriers to migration. The potential effects of the climate trend on the geographic range of arthropods are commonly evaluated by climate-matching models, a set of methods based on the recorded distribution that assess the potentially available range for a species according to its preferences for a group of explanatory variables. Process-driven models focus on each part of the life cycle, and are considered a essential tool for research of tick-borne pathogen transmission rates (Randolph and Rogers 2003). Efforts to construct process-driven models have been focused on Ixodes ricinus (Hancock et al. 2010), a conspicuous vector of Lyme borreliosis and TBE in many parts of Europe (Jaenson and Lindgren 2011).
A process-driven model was recently built for H. marginatum (Estrada-Peña et al. 2011). The model was focused not only on understanding the regulation of tick seasonality by dominant climate features, but was also designed to examine the potential effects of a changing environment on the colonization potential of the tick in areas outside its current range. This study aimed to examine the potential effects of future climate on the life cycle of the tick across the Mediterranean basin based on a set of previously built climate scenarios. We specifically sought to describe areas where climate features and a critical habitat configuration for host dispersal would result in major changes in tick turnover. We built a model based on the host movement rules over the interface of the process-driven tick model, regulated by the daily climate obtained for a spatially interpolated 10-min grid over the target region.
Materials and Methods
Sources of climate data
The CRU CL 2.0 (Mitchell et al. 2004) dataset was used for current climate data (
Scenarios for future climate conditions were obtained from the CGIAR Research Program on Climate Change, Agriculture, and Food Security (CCAFS) web site (
Process-driven models to evaluate life cycle changes
Both current and projected climate scenarios contain the data necessary to run a process-driven model for H. marginatum (as built by Estrada-Peña et al. 2011). Our aim was not to compute absolute changes in the phenology of the tick according to climate scenarios, but to obtain relative estimations. We sought to obtain the averaged effect of the different climate scenarios in the years 2020, 2050, and 2080 compared to the “normal” climate (years 1961–2000). Such an effect was summarized over a background, averaging outputs along a standard set of regions. Ecological regions were downloaded from
Methods to assess dispersal
Modeling the invasive behavior of ticks is a complex task that involves not only life cycle traits, but also other data related to the way hosts move and disperse (Estrada-Peña 2004). We evaluated sites where a high exchange rate of ticks among vegetation patches is expected (due to host movements), and where λ>0 (due to life cycle traits). We called the combination of both values the “probability of occupancy.” We first formulated host dispersal as a cellular automata, modeling their movements in a network linking patches of vegetation according to a set of general rules derived from the topology of the network, and friction surfaces against which hosts move. The number of ticks passively moving from one patch to another is estimated as the probability of hosts moving along every link between any two patches. For simplicity, it is specifically assumed herein that ticks use only one general type of host, and that host availability is a function of the patch size. This assumption was made because of the lack of adequate knowledge regarding hosts used across the geographic range of the tick, and the inherent difficulty of parameterizing preferences towards each host. Local variations in the host distribution pattern must be expected, but H. marginatum is a generalist feeder, and equilibrium of host availability should be assumed across its range.
The connectivity of heterogeneous landscapes is not easily captured by simple index or landscape metrics. We used the MODIS MOD12Q1 data set of remotely sensed information, which classifies land cover into vegetation categories. The nominal resolution of 1 km may be too coarse to capture some very local processes, but tests performed with the same imagery at 500 meters resolution rendered unacceptably high computation times. GIS software (Erdas Imagine 9.3) was fed data on dominant vegetation at 1-km resolution, the vegetation patches were vectorized, and a network linking every pair of resulting patches was produced. As a measure of potential immigration to each patch i from the total patches, patch connectivity (Si) was estimated as (Hanski 1994; Moilanen and Nieminen 2002):
Connectivity increases with the area of each patch j, but decreases with its distance from patch i. The constant α estimates how migration from patch j declines with increasing distance and is the slope of a negative exponential distribution. Dij is the distance between the centers of patches i and j; Aj is the area of patch j; and B relates emigration rate to patch area (Hanski 1994, Moilanen and Nieminen 2002). We estimated connectivity using α=2 and B=0.5, accounting for the tendency of per capita emigration to be greater from smaller habitat patches (Thomas and Hanski 1997). Thus, connectivity is a general measure of animal movement among patches according to their sizes and relative distances. High connectivity values are obtained for small, well-connected patches very near to one another, but low connectivity values indicate large or relatively small, but always isolated, patches.
Connectivity values were further weighted by friction surfaces based on the asymptotic function of elevation and slope obtained from a digital elevation model (DEM) of the target area downloaded from the GLOBE project (

The methodological framework used in this study. (
Results
Table 1 includes the net changes in λ values in the different ecoregions of the target area. Averaged λ values tend to increase along the time window for every ecoregion, more than doubling by 2080 compared to 1961–2000 estimates. Some sites where the tick is currently absent would support permanent populations in the near future. However, simulations also predicted that some areas where the tick is now present would turn into an unsuitable habitat during the same time periods. Table 2 presents the mean values of duration of oviposition and nymphal-adult molt in the different ecoregions according to the climate scenarios. Though the oviposition period tends to become slightly longer, the nymphal-adult molt tends to decrease, up to 2 weeks on average. The survival rates during oviposition and molting (Table 3) were predicted to slightly change and greatly increase (around 3 to 4 times), respectively. Some regions in the Caspian and the Caucasus are predicted to be unsuitable under current climate conditions, but would allow nymphal molt survival up to 11–14% with future climate change. That rate is seven times higher than the average survival under current conditions for the whole target area in Europe and Africa. Table 4 presents the data on survival during the questing adult stage, showing the same trend toward an increase according to the projected climate scenarios.
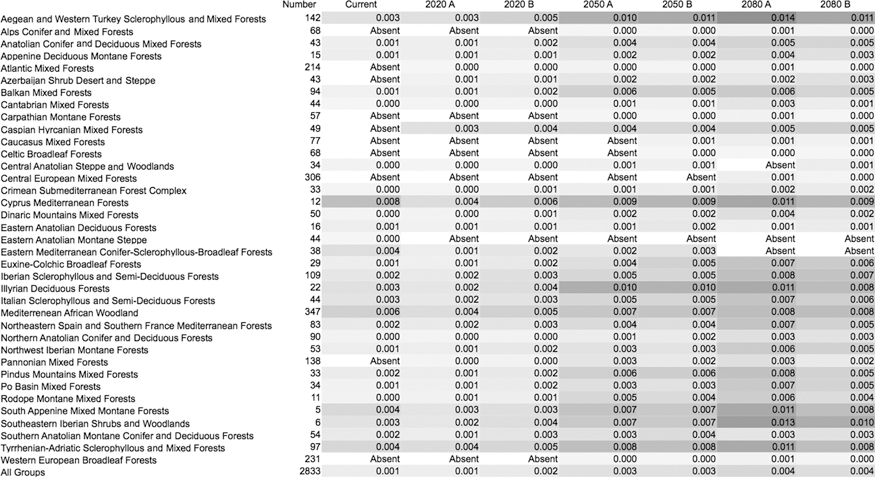
Included are the number of 10-min cells partially or totally included within each ecological assemblage. A shadow of grey has been provided for visual comparison of changing values across the different scenarios. Included are the date for “current” climate (1970–1999), and for the time slices 2020, 2050, and 2080, considering the A (high-emissions) and B (low-emissions) scenarios.
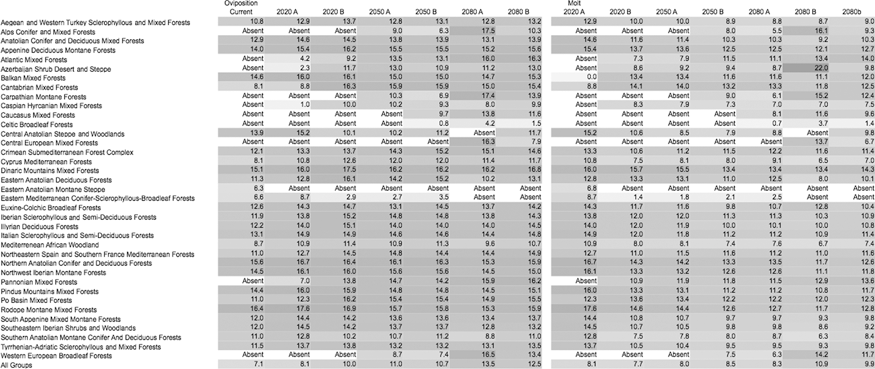
Data are in weeks and are the average of the cycle in one complete calendar year, separately for each ecological region in the target area. A shadow of grey has been provided for visual comparison of changing values across the different scenarios. Included are the date for “current” climate (1970–1999) and for the time slices 2020, 2050, and 2080, considering the A and B scenarios.
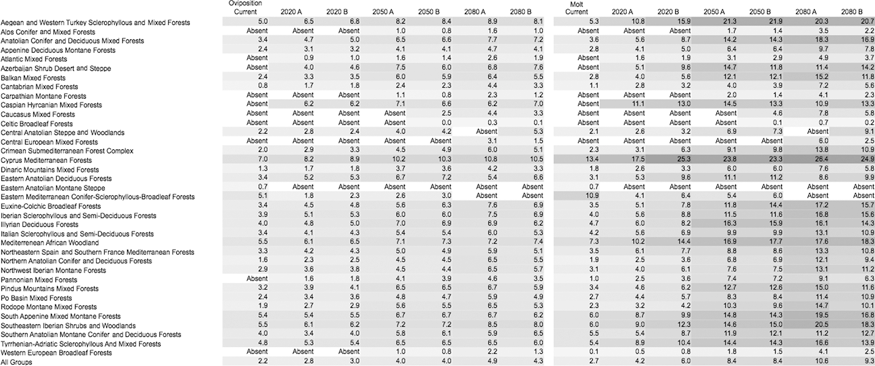
Data are in percent. A shadow of grey has been provided for visual comparison of changing values across the different scenarios. Included are the date for “current” climate (1970–1999) and for the time slices 2020, 2050, and 2080, considering the A and B scenarios. Data are provided separately for each ecological region in the target area.

Data are in percent. A shadow of grey has been provided for visual comparison of changing values across the different scenarios. Included are the date for “current” climate (1970–1999) and for the time slices 2020, 2050, and 2080, considering the A and B scenarios. Data are provided separately for each ecological region in the target area.
Figure 2 shows the changes in probability of occupancy for single cells in the honeycomb overlaying the target area. The most drastic increases in the probability of occupancy were observed over Italy, south of the Alps, the Balkans, the Carpathians (Romania, Ukraine, and Moldavia), wide areas of southern Russia (north of the Black Sea), and in some areas of Germany and the Netherlands. The probability of occupancy remained relatively unchanged in the range of current distribution experiencing the warmest current climate (e.g., western Turkey or northern Africa). The northern latitudinal range of the tick is expected to increase, providing connections among standing populations in the western Mediterranean and western Balkans that are currently highly disconnected.
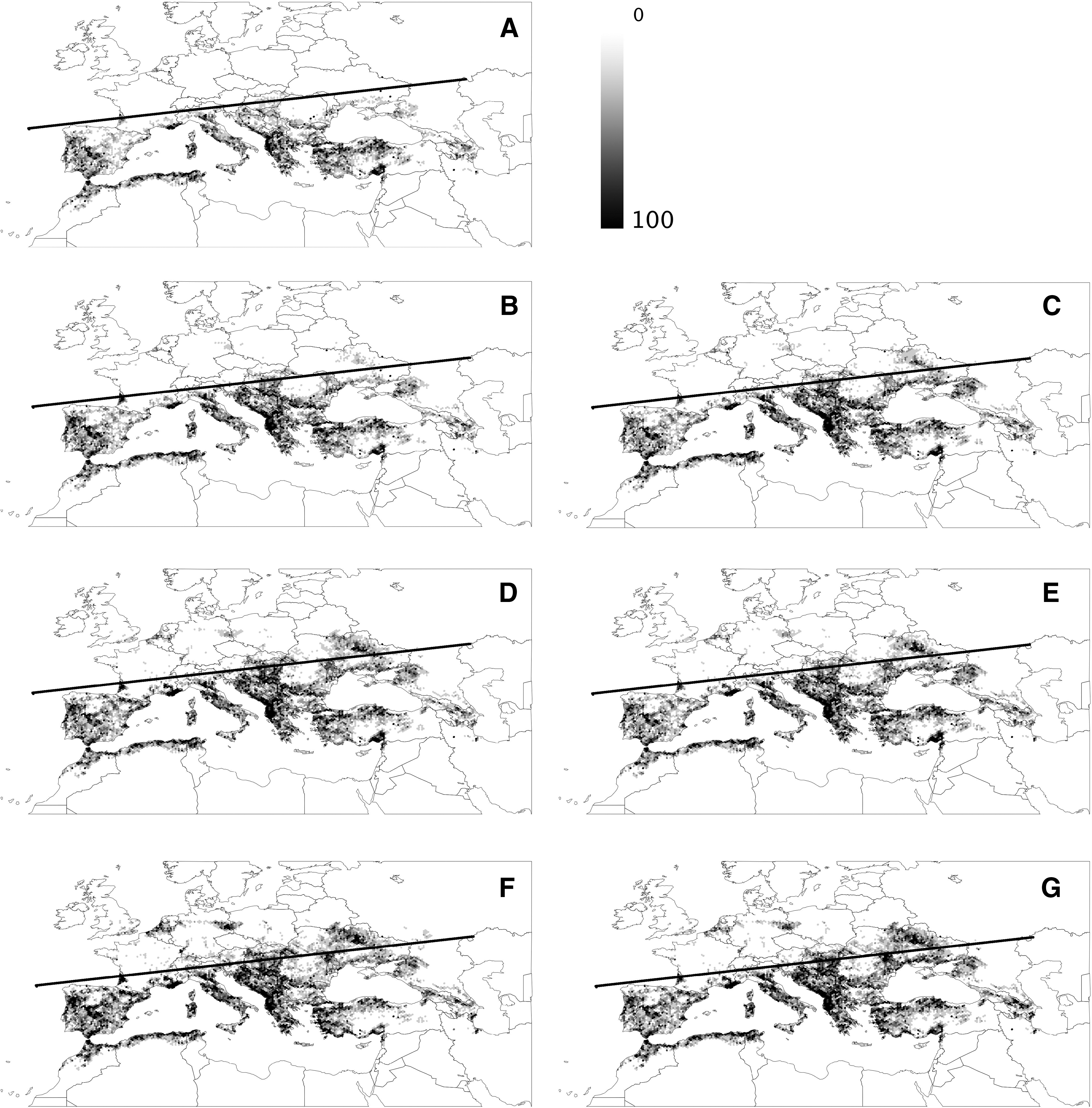
Changes in the computed probability of occupancy for H. marginatum in the western Palearctic. The measure is unitless, combining the net growth rate of the tick in the site and the connectivity of vegetation patches due to host movements. Data on probability of occupancy range from 0 to 100. (
Discussion
We demonstrated that the range of H. marginatum is expected to increase northwards in the western Palearctic because of climate trends, assuming an equilibrium between the tick and its environment. Our study is an effort to model the spread of ticks according to a series of rules that guide host movements and link to traits obtained from an already-tested process-driven model of the tick life cycle. Efforts have been made to evaluate the dispersion rates of Ixodes scapularis in the U.S. (Madhav et al. 2004; Ogden et al. 2005), and to determine changes in abiotic suitability for ticks in the Mediterranean region (Estrada-Peña and Venzal 2007). H. marginatum is an arthropod threatening animal and human health and the subject of current research because of its ability to transmit the CCHF virus, which causes a serious human disease. Gale and colleagues (2009) already outlined the feasibility of developing a risk assessment from a series of informative layers describing the risk of the spread of CCHF into Europe. Future climate has been speculated to improve the survival of the tick, and will allow northern colonization of tick populations because of the introduction of specimens by birds, or the spread of populations living near the environmental limits of effective colonization, close to the lower thermal thresholds that are likely to be exceeded in the next few decades (Gale et al. 2009).
The process-driven model explicitly assumed that the modeled organism cannot adjust its life cycle to fit shifts in environmental conditions. However, this point probably does not match the real performance of organisms during climate change, and they will likely acclimate for maximum performance under the new conditions. Unexpected results were observed for the development and mortality rates from oviposition to larval hatching, and during the nymphal-adult molt. Development rates were slightly slower, whereas the survival during the nymphal-adult molt was clearly greater. The former result seems to be due to the daily temperatures exceeding the optimal threshold for the tick, whereas the latter is due to the impact of a warmer climate on the survival ability of a thermophilic tick (Morel 1965). The average survival rates of questing adults are also higher in future climate scenarios. The conclusion for the climate scenarios is that λ, a comparative measure of life cycle performance, will increase in most ecoregions of the western Palearctic.
Concerns exist about the possible spread of the CCHF virus north and west of the current distribution of the tick that carries it, but evaluating how vector and reservoir populations will change in a warmer climate is necessary before confirming these concerns. Cases of disease are currently reported only in parts of the Balkans, Turkey, and Russia (Ozkaya et al. 2010; Ozdarendeli et al. 2010; EFSA 2010). One of the possible reasons for such an apparent absence would be the possible lack of connectivity between eastern (infected) and western tick populations (presumably free of more pathogenic strains of the virus). Viral strains are different in eastern and western Thrace (between Greece and Turkey), due to a lack of mixing of the virus over its long history (Mild et al. 2010). If H. marginatum in southwestern Europe is distributed in relative isolation from neighbor populations, the genetic flow of the populations will show specific signatures of isolated specimens, which should be examined.
With warmer scenarios, new areas become available for tick survival, enabling the spread of ticks north and west of the Balkans. These results must also be interpreted with caution; they are intended as a preliminary assessment of the impact of climate on tick survival rates. This assessment did not include the effects on host abundance and response to habitat, or vegetation traits, and must be taken as a preliminary evaluation. H. marginatum is passively dispersed by numerous host species with differing movement abilities, with different tick stages often having different host preferences (Apanaskevich and Horak 2008). Even with the restrictive conditions of our modeling approach, the conclusion is that the only barrier to tick spread (and probably pathogenic CCHF virus strains) is the minimum annual temperature. The northern limit would expand into northern latitudes, but the southern limit would not change substantially.
Our results offer tantalizing future directions to study complex tick dispersal mechanisms. The coupling of a process-driven model with a simple model for host movements based on the real landscape provided an interesting framework to assess probable routes of tick dispersal based on differing climate scenarios. Ultimately, a spatially explicit model should allow us to understand the effects of complex movement routes on the emergence of risk for tick-borne diseases. The incorporation of more realistic host effects (e.g., different types of hosts, different movements, and seasonality of behavior), coupled with the power of the process-driven model for the tick life cycle, will improve our knowledge of the dynamics of the tick-virus system, improving management measures against disease.
Footnotes
Acknowledgments
This study was conceived and designed in an international meeting in Istanbul, Turkey, in 2007, and aimed to elucidate the basics of the apparent spread of the CCHF virus, organized by the European Integrated Consortium ICTTD3. The authors want to acknowledge preliminary discussions with the people attending the meeting. We also gratefully acknowledge the contributions of those who contributed to the main dataset of tick records, and to our approach to understanding the ecology of H. marginatum. Special thanks are due to the support and framework provided by ArboZoonet, an International Network for Capacity Building for the Control of Emerging Viral Vector-Borne Zoonotic Diseases, project number 211757, under the European Union FP7 framework.
Author Disclosure Statement
No competing financial interests exist.