
Abstract
The recent range expansion of Ixodes scapularis has been accompanied by the emergence of Borrelia burgdorferi. The development of genetic diversity in B. burgdorferi at these sites of emergence and its relationship to range expansion is poorly understood. We followed colonization of I. scapularis on a coastal Maine island over a 17-year period. B. burgdorferi's emergence was documented, as was expansion of ospC strain diversity. Ticks collected from rodents and vegetation were examined for the presence of B. burgdorferi. Sequencing and reverse line blot were used to detect B. burgdorferi ospC major groups (oMG). No I. scapularis were found until year four of the study, after which time they increased in abundance. No B. burgdorferi was detected by darkfield microscopy in I. scapularis until 10 years into the study, when 4% of adult ticks were infected. Seven years later, 43% of adult ticks were infected. In 2003, one oMG accounted for 91% of B. burgdorferi strains. This “founder” strain persisted in 2005, but by 2007 was a minority of the 7 oMGs present. Given the island's isolation, gene flow by avian introduction of multiple strains is suggested in the development of B. burgdorferi oMG diversity.
Introduction
The establishment and spread of I. scapularis populations is relatively recent (Spielman et al. 1985; White et al. 1991; Rand et al. 2007), and is driven by changing land use and vegetation patterns and increasing abundance of white-tailed deer (Odocoileus virginianus) (Ginsberg 1993; Piesman 2002). Dispersal of I. scapularis by both birds (Smith et al. 1996; Scott et al. 2001; Brinkerhoff et al. 2011) and deer (Bouseman et al. 1990) are postulated to contribute to its range expansion.
Once the vector population has been established, an epizootic B. burgdorferi cycle can be perpetuated. This may be accomplished when the spirochete is dispersed to a new area by infected reservoir hosts, such as birds (Brinkerhoff et al. 2010), or by larval deer ticks that are attached to infected small mammals or birds (Rand et al. 1998; Ginsberg et al. 2005; Brinkerhoff al. 2011). In the northeastern United States, both the tick and spirochete are now well established across a broad region (Dennis et al. 1998), including many areas of northern New England (Anderson et al. 1987; Ginsberg and Ewing 1988; Diuk-Wasser et al. 2006). In other areas, Lyme disease is still emergent, with few cases and sporadic numbers of ticks found each year (Rand et al. 2007).
Recent studies suggest that B. burgdorferi strain diversity may contribute to enzootic maintenance of B. burgdorferi populations (Brisson and Dykhuizen 2004). Some small mammals, particularly Peromyscus mice, sciurids, and shrews are more efficient hosts and transmitters of certain strains of B. burgdorferi than others (Brisson and Dykhuizen 2004). At sites with a limited number of small mammal host species, the presence of a full complement of B. burgdorferi strains could increase the likelihood that the population will be maintained. Within established populations of B. burgdorferi, there is locally high genetic diversity that may be maintained by frequency-dependent balancing selection (Wang et al. 1999). This diversity appears to be universal in the northeast United States (Qiu et al. 2002), including at sites on mainland coastal Maine (Mathers et al. 2005). However, the mechanism of the establishment of B. burgdorferi diversity in emergent areas is unknown.
Outer surface protein C (ospC) allele diversity can be used to describe genetic diversity in B. burgdorferi (Wang et al. 1999; Brisson and Dykhuizen 2004). ospC codes for a 23-kDa protein that allows B. burgdorferi to enter I. scapularis salivary glands while the tick is feeding (Pal et al. 2004). There are 21 designated ospC major groups (oMG) (A through U), though others remain undescribed. By definition, each group is made up of alleles of the ospC gene with nucleotide sequences that are on average more than 99% similar (Wang et al. 1999).
Previous studies of B. burgdorferi strain diversity have been carried out in areas of established B. burgdorferi populations (Wang et al. 1999; Qiu et al. 2002). Our work documents the establishment of I. scapularis populations followed by the emergence of B. burgdorferi on an island on the coast of Maine where neither had previously been found (Rand et al. 1991; Smith et al. 1992). In this report, we include the unique description of the evolution of B. burgdorferi strain diversity within a newly established population of hard ticks in the isolated setting of island ecology.
Materials and Methods
Specimen collection
Swan's Island is a 36.1 km2 island located in the Gulf of Maine (44.15o N/68.44o W) 7 kilometers from the coast of Maine (Fig. 1). The island habitats consist of coniferous and mixed forests and areas of dense shrub. Fields and lawns are present around year-round and seasonal houses. White-tailed deer are present, as are domestic cats (Felis sylvestris) and dogs (Canis familiaris), deer mice, meadow voles (Microtus pennsylvanicus), red backed voles (Myodes gapperi), masked shrews (Sorex cinereus), raccoons (Procyon lotor), European hares (Lepus europaeus), muskrats (Ondatra zibethicus), red squirrels (Tamiasciurus hudsonicus) and mink (Mustela vison).

Map of coastal Maine, including Swan's Island.
We conducted flagging surveys for questing adult ticks at their seasonal peak (Rand et al. 1998) for Maine: mid-October through mid-November. Adult ticks were chosen due to their ease of collection compared to immature ticks. Field surveys were conducted in 1990, 1994, 1996, 1997, 2000, 2001, 2003, 2005, and 2007. We flagged vegetation with a 1-m2 flannel cloth with checks for ticks every 15 sec (Markowski et al. 1997) between 0900 and 1600 h on days when the temperature was above 10°C.
Live trapping of small mammals was also undertaken, starting in 1990, and then in 1995, 1996, 1997, 2003, and 2005. At the beginning of each survey, we placed a Sherman live trap (model #3310A; HB Sherman Co., Tallahassee FL) in shaded habitats conducive to ticks (Ginsberg and Ewing 1988). Upon capture, ectoparasites were collected (Keirans and Litwak 1989; Durden 1996). Ear snips were also taken from live mice using sterile technique and placed in liquid Barbour-Stoenner-Kelly (BSK) media (Sinsky and Piesman 1989). Captures were subsequently released. Field surveys of small mammals were performed in accordance with protocols approved by the Institutional Animal Care and Use Committee at the Maine Medical Center Research Institute (protocol #1005).
Detection of Borrelia burgdorferi
Live I. scapularis adults, stored at 4°C since collection, were analyzed by direct fluorescent antibody (DFA) for the presence of B. burgdorferi in 1994, 1996, 1997, 2000, 2001, 2005, and 2007. They were dissected and their midguts examined for the presence of B. burgdorferi by fluorescence with a polyclonal anti-Borrelia antibody (Donahue et al. 1987). Each tick was considered positive if 3 or more spirochetes were seen with 400×epifluorescence microscopy.
In 2007, 125 ticks also were tested for B. burgdorferi DNA by polymerase chain reaction (PCR). After collection, ticks that were to be analyzed by PCR were placed in individual vials and stored at −70°C. Tick and spirochete genomic DNA was extracted using a Qiagen DNeasy Blood & Tissue Kit (Qiagen, Valencia, CA). The ospC region of B. burgdorferi was amplified by PCR using a Qiagen HotStarTaq Master Mix kit (Qiagen). The primers were OC+: 5′AAA GAA TAC ATT AAG TGC GAT ATT 3′ and OC−: 5′GGG CTT GTA AGC TCT TTA ACT 3′, and yielded an approximately 600-base-pair product if positive (Wang et al. 1999). PCR conditions were initial activation at 95° for 15 min followed by 35 cycles of denaturation at 94° for 1 min, annealing at 52° for 1 min, and extension at 72° for 1 min, followed by a final extension at 72° for 10 min.
To determine whether there was a significant trend over time in both ticks collected per hour and B. burgdorferi infection rate, we log-transformed the responses and modeled each as a function of time.
Determination of ospC genotype in 2003 and 2005
In 2003, 2005, and 2007, the B. burgdorferi ospC genotype in each tick was determined. In 2003 and 2005 this was accomplished by sequencing the ospC region of DNA extracted from B. burgdorferi cultures. Midguts dissected from ticks and mouse ear snips were placed in individual vials of BSK media and incubated at 34°C. After 2 weeks they were examined by darkfield microscopy for the presence of spirochetes. Genomic DNA was extracted from positive cultures with the Qiagen MiniPrep kit (Qiagen). The ospC region of B. burgdorferi was amplified by PCR as described above. The PCR product was purified with a Qiagen QIAquick PCR Purification kit (Qiagen). Samples were sequenced by a standard protocol using a rhodamine ready reaction kit (Applied Biosystems, Foster City, CA) with an automated (ABI 310) sequencer.
Sequences were aligned using Clustal X (Thompson et al. 1997), then adjusted by eye using Se-Al software (Rambaut 2002). Ambiguous sequences were repeated. If after re-sequencing the chromatogram remained ambiguous, the sequence was excluded from further analysis.
Determination of ospC genotype in 2007
To detect multiple B. burgdorferi oMGs in each tick, and because of transient loss of sterility in the Borrelia culture system utilized in previous years, reverse line blot (RLB) was used to analyze each positive tick in 2007 (Saiki et al. 1989; Qiu et al. 2002; Brisson and Dykhuizen 2004). In RLB, a nucleotide probe specific for each oMG is anchored to a membrane and binds its complementary sequence in a DNA sample. The DNA in this case was a PCR product amplified from genomic B. burgdorferi DNA with a biotin label on the 5′ end of primer “OC+,” which allowed use of a streptavidin-horseradish peroxidase system to detect binding (Roche, Mannheim, Germany). Probes to ospC DNA specific to each oMG, except C, were used for each sample. oMG C is a recombinant of groups B, I, and E, and no specific probe is available for it, so it could not be detected. To ensure the reproducibility of RLB as compared to sequencing, six samples of surplus genomic DNA from 2005 that had been extracted from cultured spirochetes and preserved at −70°C were re-examined with RLB.
Results
During flagging in 1990, no I. scapularis were found. In 1994, 0.1 adult I. scapularis was flagged per hour. By 2007, 8.7 adults were flagged per hour (Table 1), a significant increase over time (n=7, p=0.0002, R2=0.95).
DFA testing for B. burgdorferi in flagged adult ticks started in 1994, but spirochetes were not detected until 2000. Infection prevalence among deer ticks increased from 4% in 2000 to 43% in 2007 (Table 1). Forty-four percent of I. scapularis flagged in 2007 were positive by PCR. The increase in infection rate over time was significant (n=7, p=0.0002, R2=0.95).
A number of rodents were trapped during field work at Swan's Island; predominantly P. maniculatus and smaller numbers of M. pennsylvanicus and T. hudsonicus. No ticks were found on trapped rodents until 1995, when I. scapularis larvae and nymphs were removed from rodents (Table 2).
In 2003, 14 P. maniculatus were trapped and 28% of their ear snip cultures grew B. burgdorferi. Twenty-two I. scapularis larvae and 4 nymphs were collected from the rodents; 35% of them contained B. burgdorferi by DFA. In 2005, 57 P. maniculatus were trapped and 32 ear snips were taken. Thirty of the ear snips were cultured and 8 (27%) were positive for B. burgdorferi.
Analysis of strain types revealed that oMG H accounted for 91% of oMGs in samples from both flagged ticks and mouse tissue culture in 2003 (Fig. 2). Four of 4 B. burgdorferi-positive ear snips contained oMG H, as did 6 of 7 adult I. scapularis. The seventh tick contained oMG A. In 2005, oMG H represented 65% of oMGs and was still the most common strain found in both mice and captured ticks. However, that year oMGs A, B, and K were isolated from rodent ear snips, while F and A were found in flagged ticks. In 2007, 7 B. burgdorferi oMGs were detected by RLB. That year there was an increase in the proportion of groups A, K, M, N, and U, while oMG H accounted for only 1% of oMGs. In 2007, the mean number of oMGs per tick was 1.74. This number could not be calculated previously because multiple oMGs could not be detected with the techniques used to measure strain diversity at that time.
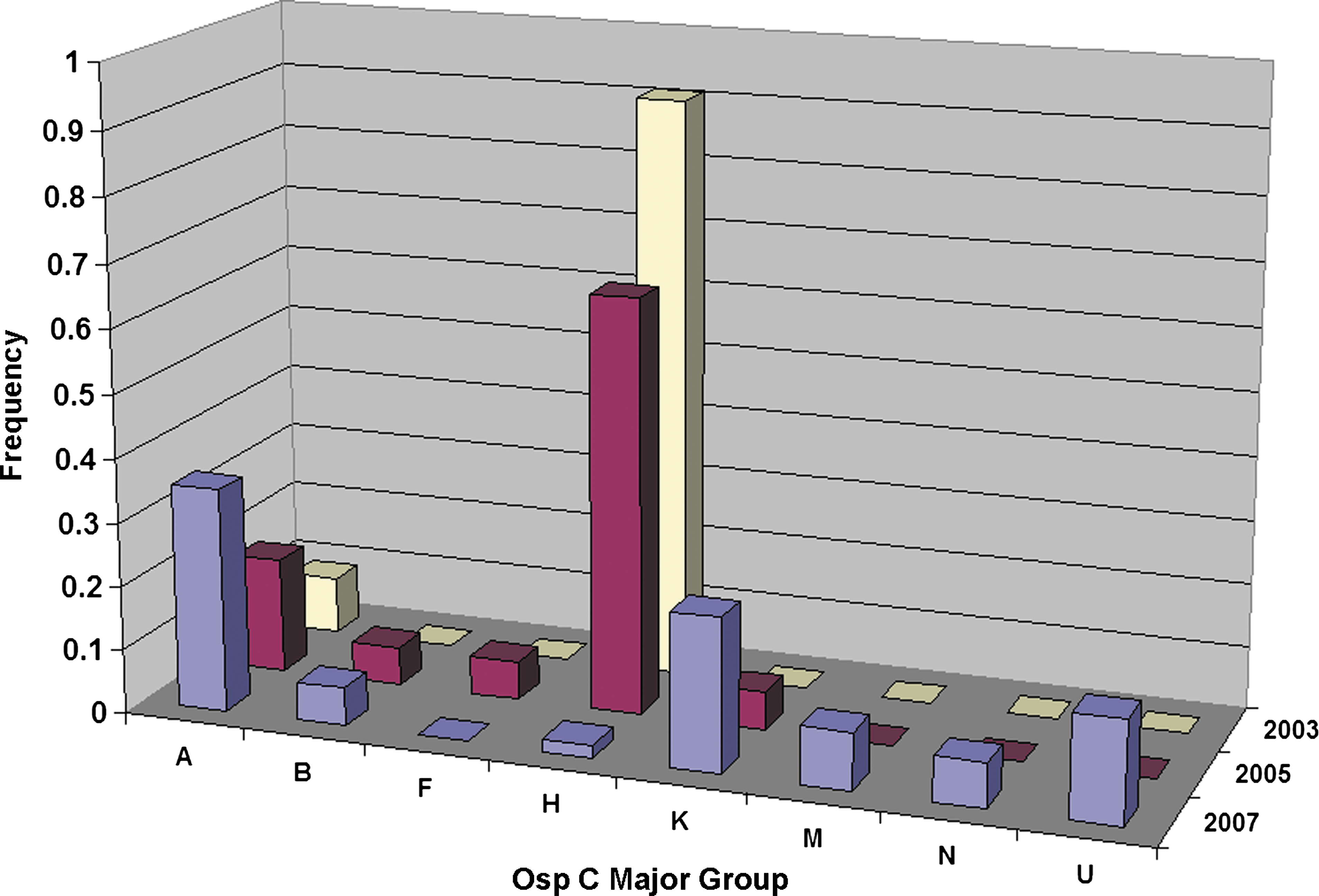
Frequency of ospC major groups (oMGs) found in ticks and rodents collected at Swan's Island, Maine over time.
RLB performed on six samples that were sequenced in 2005 to determine genotype were found to contain oMGs K, H, B, H, H, and A by each method.
Discussion
I. scapularis and B. burgdorferi populations were established sequentially on a coastal Maine island and became more abundant over time. In 1990 there were no ticks found during an initial survey, but within 5 years they were detected in low numbers, both while flagging and on small mammal hosts. They continued to increase in abundance through 2007. This observation supports theoretical models that predict a transient low density of I. scapularis before colonization of an area (Madhav et al. 2004). It also provides evidence for a timeline of establishment of reproducing I. scapularis populations over years to decades (Wilson 1998).
Despite sampling ticks and rodents since 1994, B. burgdorferi was not found until 2000. This suggests a lag time between I. scapularis colonization and emergence of an enzootic B. burgdorferi cycle. The rare I. scapularis specimens that were initially collected may have been introduced arthropods that pre-dated a reproducing population. The lag between tick and B. burgdorferi emergence may represent the time needed to establish reproducing I. scapularis populations.
As demonstrated in other studies of Lyme emergence, when B. burgdorferi was eventually found in ticks, it was present at low frequency. Northward migrating birds, either by carrying immature infected ticks, or by serving as a reservoir themselves for B. burgdorferi (Brinkerhoff et al. 2010), are the most likely means of spirochete introduction (Dennis et al. 1998). At other sites, B. burgdorferi incidence in newly colonized I. scapularis populations mirrored infection rates in ticks removed from birds. This was followed by a decline in B. burgdorferi prevalence due to dilution of the spirochete in the local population of uninfected ticks (Ogden et al. 2010). The rapid increase in B. burgdorferi prevalence we observed suggests that the low prevalence observed in those tick populations may be short lived. Our results are also in accordance with a previously described model of the spread of B. burgdorferi in I. scapularis and mammal hosts. It predicted low levels of B. burgdorferi infection for a number of years after introduction, followed by rapid increases in B. burgdorferi prevalence in the tick population (Ginsberg 1988).
Once reproducing I. scapularis populations are established, arrival of infected ticks or reservoir hosts allows the Lyme disease spirochete to create an enzootic cycle at an emerging site (Matuschka and Spielman 1986). The order of tick and spirochete emergence at Swan's Island is consistent with the “tick first” hypothesis suggested as one of three scenarios for the emergence of B. burgdorferi and its vector (Hamer et al. 2010). Hamer suggests that deer are responsible for introducing uninfected ticks to uninfested areas. Large mammals such as deer can disperse ticks over short distances (Bouseman et al. 1990; Wilson 1998; Madhav et al. 2004). At our study island, this is less likely because there is a higher barrier to deer migration than at a mainland site. Instead, uninfected larval ticks were likely introduced to the island by migrating birds. This allowed for a unique evaluation of the emergence of B. burgdorferi with avian introduction as the predominant variable. Long distance dispersal of engorged larvae and nymphs to isolated locations by migratory birds has been documented (Smith et al. 1996; Scott et al. 2001; Mathers et al. 2011; Elias et al. 2011). The potential for spirochetemic migrating birds themselves to serve as the original B. burgdorferi reservoir has also been shown and is likely at this northeastern island site (Rand et al. 1998; Ginsberg et al. 2005; Brinkerhoff et al. 2010).
Initial studies of B. burgdorferi strain diversity (2003–2005) revealed a predominant strain (oMG H) on the island. By 2007, however, there was loss of this initial “founder” strain and gradual replacement with a complement of strains that more closely resembles other mainland sites with established B. burgdorferi populations (Mathers et al. 2005). Notably, oMG A became the most common strain, which has been seen in surveys at other sites in the Northeast, but not as commonly in the Midwest (Qiu et al. 2002; Gatewood et al. 2009). Previous studies have not documented this changing molecular epidemiology through time at sites of emerging infection.
The increase in I. scapularis abundance in our study was followed closely by an increase in B. burgdorferi infection rate and oMG diversity. It is possible that higher oMG diversity improved B. burgdorferi transmission among ticks and small animal reservoirs, and thereby improved its survival. It is also likely that threshold densities of both ticks and spirochetes must be reached to improve the efficiency of the zoonotic cycle. In areas of New Hampshire with the highest tick densities, higher B. burgdorferi infection rates in I. scapularis have been found (Walk et al. 2009).
Previous authors have attributed local changes in oMG prevalence to frequency dependent selection by secondary responses of host immune systems (Wang et al. 1999). A model of “multiple niche polymorphism,” a type of balancing selection, has been proposed to explain changes in oMG frequency. In this model, certain strains can only infect a subset of hosts. As such, host population dynamics have an impact on B. burgdorferi strain diversity. The maintenance of high oMG diversity is postulated to allow B. burgdorferi to survive in an environment that can include relative changes in host species abundance (Wang et al. 1999; Brisson and Dykhuizen 2004).
In order to balance selection to maintain high diversity, the subset of existing oMGs that it acts on must be diverse to begin with, even if some strains are present only at low frequencies. At a site of emerging infection, we found low initial oMG diversity, which does not provide a substrate for selection. The most likely explanation for the observed increase in diversity there is by gene flow from infected migrating birds or engorged larval ticks on them. Previous work at another coastal Maine site showed a diverse complement of oMGs in ticks removed from migrating birds (Mathers et al. 2011). If migration is occurring at sites of emerging infection, and there is no barrier to it in populations that are well established, it may also play a role in maintaining high genetic diversity in enzootic populations.
Low initial oMG diversity may have been due to sampling bias during field collections. However, our sample of ticks and mammals represented diverse field sites. Low diversity could have been due to variations in oMG detection techniques. It was measured by sequencing DNA extracted from cultured B. burgdorferi in 2003 and 2005, and by RLB of DNA extracted from ticks in 2007. While the two systems gave reproducible results, the use of culture may have restricted the number of oMGs detected.
Our work provides insight into the geo-temporal trends seen in Lyme disease emergence and suggests gene flow as a factor in maintaining its genotypic diversity. It outlines a time course for emergence and establishment of the disease, which has implications for assessing the risk of exposure for human and animal populations. Future work is needed to determine if emergence of other tick-borne infections fits this framework, or if it is unique to Lyme disease.
Footnotes
Acknowledgments
Thanks to the laboratory of Dr. Calvin Vary for assistance with nucleotide sequencing. This study was funded by a grant from the Maine Medical Center Research Institute (MMCRI).
Author Disclosure Statement
No competing financial interests exist.