
Abstract
Human cutaneous leishmaniasis is one of the most challenging public health issues in many tropical and subtropical countries of the world, including Iran. More than half (54%) of the new zoonotic cutaneous leishmaniasis (ZCL) cases among the Eastern Mediterranean countries were reported from Iran in 2008. The detection of Leishmania parasites in rodents is essential to incriminate them as probable reservoir hosts of ZCL infection. As a result of the annual detection of about 200–250 clinical ZCL cases in the Jask district of southern Iran, feral rodents were trapped, identified to species level, and examined for Leishmania presence by preparing routine blood smears on microscopic slides from 2007 to 2008. Overall, 27 Tatera indica, 17 Gerbillus nanus, 29 Meriones persicus, 26 M. hurrianae, and 7 M. libycus were identified. Females of T. indica, M. hurrianae, and G. nanus appeared to be naturally infected with the protozoan parasite, L. major. This is the first report of microscopic and molecular detection of this trypanosomatid parasite infecting these three rodents reported from Hormozgan province in southeast Iran. More than three-quarters (82%) of the parasite-infected rodents came from the eastern plain of this province, but none of the other rodents were found to be smear-positive or kinetoplast DNA-positive by PCR. M. hurrianae, G. nanus, and T. indica are therefore incriminated as three potential reservoir hosts of L. major in Oriental parts of Iran.
Introduction
Human cutaneous leishmaniasis (CL) is a sand fly-borne skin infection caused by the blood parasite Leishmania (Ashford 2000). It is still a major public health issue in Iran (with 26,824 new CL cases in 2008), where most CL cases are zoonotic, with rodents acting as reservoir hosts in many rural areas (Ruiz Postigo 2010). Zoonotic CL (ZCL) is essentially an infection of rodents with natural focality (Molyneux and Ashford 1983). In Iran, the causative agent of ZCL, L. major, is often transmitted to humans by the bites of the female sand fly, Phlebotomus papatasi, mostly from rodent hosts that act as reservoirs for the human infection (Fakoorziba et al. 2011). ZCL occurs in most rural areas of the arid and semi-arid regions in all but the northwest provinces of Iran. P. papatasi is definitely the main vector and various feral rodents, particularly but not exclusively those in the subfamily Gerbillinae (Rodentia: Muridae), are reservoir hosts (Pourmohammadi et al. 2008).
Several genera of gerbilline rodents are focally distributed within the Palearctic (Rhombomys opimus) or the Oriental regions (Tatera indica and Meriones hurrianae) of Iran. In contrast, a few gerbillines (like the Libyan jird, M. libycus) are ubiquitously distributed all over the country. These distributions usually shared with the pathogenic landscape topography of ZCL rodent reservoir hosts, describe three main regional foci of infestation (Lambin et al. 2010; Nadim 2000). The first most important focus in the central and northeast region involves the great gerbil, R. opimus, as the main stable reservoir host of hyperendemic ZCL. In the absence of the great gerbil, the Libyan jird is the main unstable reservoir host of ZCL (Gramiccia and Gradoni 2005). The second focus is represented by the Indian gerbil, T. indica, in the midwest and southern regions. The third is the southeast Oriental region involving the Indian desert jird, M. hurrianae. These are often sympatric with species in the genus Gerbillus, whose role in the eco-epidemiology of ZCL has thus far been largely neglected.
As part of a literature survey in this study, a total of nine separate species in seven distinct genera of murid rodents were enlisted to be considered as potential reservoirs of L. major in Iran (Table 1). More than one-tenth (14%) of these mainly feral and/or synanthropic rodent species could thus potentially act as reservoir hosts of ZCL pathogens.
Bandar Jask is about 330 km to the east of Bandar Abbas, the capital city of Hormozgan province, southern Iran. The numbers of ZCL cases in Bandar Jask were 223 and 245 in 2006 and 2007, respectively. Considering the province's pathogenic landscape topography and the patchy dispersion of infectious reservoirs, it is possible that the infection spreads to other similar nearby regions. This study was thus undertaken in order to identify the rodent fauna and to determine the potential reservoir hosts of ZCL in this new focus.
Materials and Methods
Study area
Hormozgan province is in the southern littoral part of Iran. It is divided into 10 districts. The most eastern one, known as Jask district (≈154 km2), is a wide aeolic plain on the northern coastline of the Oman Sea (25°38'N, 57°46'E, at an altitude of 4.8 m a.s.l.) located on the southernmost section of the Iranian mainland at an ecotone between sand dunes, with an approximately 30-km littoral plain belt to the south, and loess plains leading to the rocky mountains in the north (Fig. 1). It has long dry summers with a hot humid climate over most of its arid and semi-arid regions with sandy hills. Its mean annual temperature and precipitation are 27°C and 125 mm, respectively. This region is mainly covered with Tamarix tetrandra (Violales: Tamaricaceae) plants, which grow in salt marshes near the seasonal drainage ditches. Other halophytic plant communities include: Prosopis juliflora, Salvadora persica, Glycyrrhiza glabra, and Ziziphus spinichristi. This study was carried out from April 2007 to April 2008, around seven villages in the rural district of Jask.
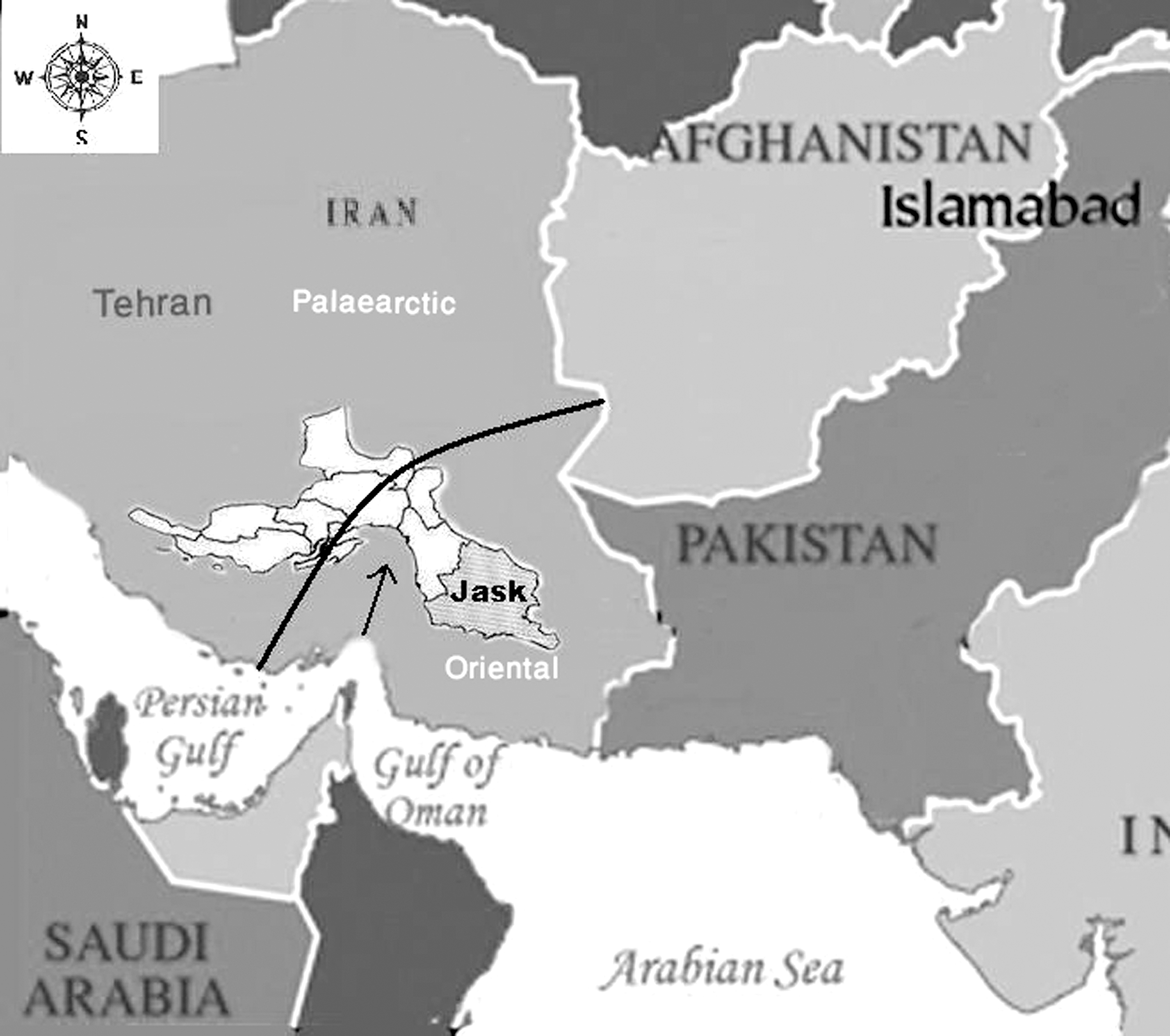
A map of Iran, showing the provincial boundaries and the location of the Jask study region.
Sample collection and rodent identification
Sample collections were conducted in spring (April–June, 2007), autumn (October–December, 2007), and winter (January–March, 2008) in the Jask district. All colonies of feral rodents were examined to locate the most usable burrows as identified by the external activity marks (loose soil on burrow holes, footprints, fresh food debris, and feces). Colonies of gerbilline rodents were almost 1.5 km away from the selected villages. Non-commensal rodents were caught once each month using 20 standard Sherman live traps (30×15×15-cm wire mesh cage traps) located next to burrow pores, and baited with a mixture of cucumbers, dates, and millet seeds. The traps were set after sunset and searched for rodents early every morning. Each live trap placed on a night was counted a “trap-night,” and there were 20 trap-nights/month.
The captured gerbils were taken to the laboratory at Bandar Abbas School of Health (HUMS), Hormozgan, Iran. Using valid taxonomic keys, they were identified to species level by morphological features after being anesthetized with chloroform (Wilson and Reeder 2005). The handling of rodents was performed under the guidelines of the Natural Resources Authority, Hormozgan Environmental Protection Organization. Preparation of smears was done following removal of their internal organs (liver and spleen), and subsequent fixation in 70% ethanol. Duplicate impression smears were taken from the tail base, ear pinnae, liver, spleen, and any patent skin lesions of every rodent, air-dried, Giemsa-stained, and checked under a compound light microscope at 1000× magnification for detection of Leishmania amastigotes.
DNA extraction
The DNA in each impression smear was extracted by digestion in lysis buffer (Motazedian et al. 2002). The dry smear was scraped off the slide using a sterile scalpel into an Eppendorf tube containing 200 μL buffer (50 mM Tris-HCl [pH 7.6], 1 mM EDTA [pH 8.0], and 1% Tween 20) with 15 μL of a solution that contained 19 mg proteinase K/mL. The tube was stirred well and incubated for 2 h at 55°C, before being centrifuged (10,000 g) for 10 min. The resultant lysate was extracted twice with phenol/chloroform/isoamyl alcohol, before the DNA was precipitated with ethanol, resuspended in 100 μL ddH2O, and desiccated to remove the ethanol. Five-microliter portions of DNA extracts were employed in PCR amplification. Each DNA suspension was stored at −20°C until use for PCR.
PCR protocol
The variable segment on mini-circles of kinetoplast DNA from leishmanial parasites was amplified using nested PCR, as described elsewhere (Azizi et al. 2011). Reference strains of L. major (MHOM/IR/54/LV39), L. infantum (MCAN/IR/96/LON49), and L. tropica (MHOM/IR/89/ARD2) were used as standards.
Agarose-gel electrophoresis
Each second-round PCR product (a 5-μL sample) was subjected to electrophoresis in 1.5% agarose gel. Using 1% ethidium bromide, the DNA bands were stained, observed with a UV transilluminator, and compared with a 1000-bp molecular-weight ladder and the relevant second-round products for the L. major, L. infantum, and L. tropica standards.
Results
Overall, 106 feral rodents were collected with an almost equal sex ratio (Table 2). They represented three genera from which five separate rodent species were identified (three Meriones, one Tatera, and one Gerbillus species). From these, one genus (Meriones) was considered common since it was represented by 62 (58.5%) individuals. No commensal rodents were trapped. The most abundant species were M. persicus (n=29, 27.4%) and T. indica (n=27, 25.5%), mainly caught from the western plain, while M. hurrianae and G. nanus (ranking the third and fourth in abundance) were largely captured from the eastern plain of the Jask region. The least abundant rodent (the red-tailed Libyan jird: M. libycus, n=7, 6.6%) was sympatric with the most abundant one, the Persian jird, M. persicus. Only two individual rodents (T. indica) were found in the central hilly areas of Gowan, while the most rodent-infested area was around Old Jask (n=23, 22%).
Although 36 gerbillines (29 M. persicus and 7 M. libycus) were also caught and checked for infection, amastigotes were only detected in smears of five (≈7%) of the 70 gerbilline rodents (M. hurrianae, G. nanus and T. indica) caught. Since infected specimens of M. hurrianae, T. indica, and one G. nanus were found in female rodents, a gender-biased parasitism with L. major was suspected. In order to confirm this, a statistical analysis was performed for significance. A simple Fisher's exact test for cumulative (i.e., only infected species) male (1/37) versus female infection rate (4/33) showed no significant difference in proportion (p>0.05).
Rodents from the eastern village of Lirdaf showed the highest positivity of infection (n=3, 15%) with L. major (Fig. 2). The highest positivity (n=2, 11.8%) of rodents confirmed molecularly to be infected with L. major was attributed to the Baluchistan gerbil, G. nanus. The eastern plain of the Jask district had more than three-quarters (82%) of the Leishmania-infected rodents. Although the number of various rodent species caught from the western plain was approximately 1.5 times that of the eastern plain, the number of L. major-positive rodents was fourfold in the latter, as it lies next to the Sistan-Baluchistan province bordering Pakistan's littoral region. Infective forms of L. major parasites were seen in each of the duplicate impression smears from each infected gerbilline rodent.
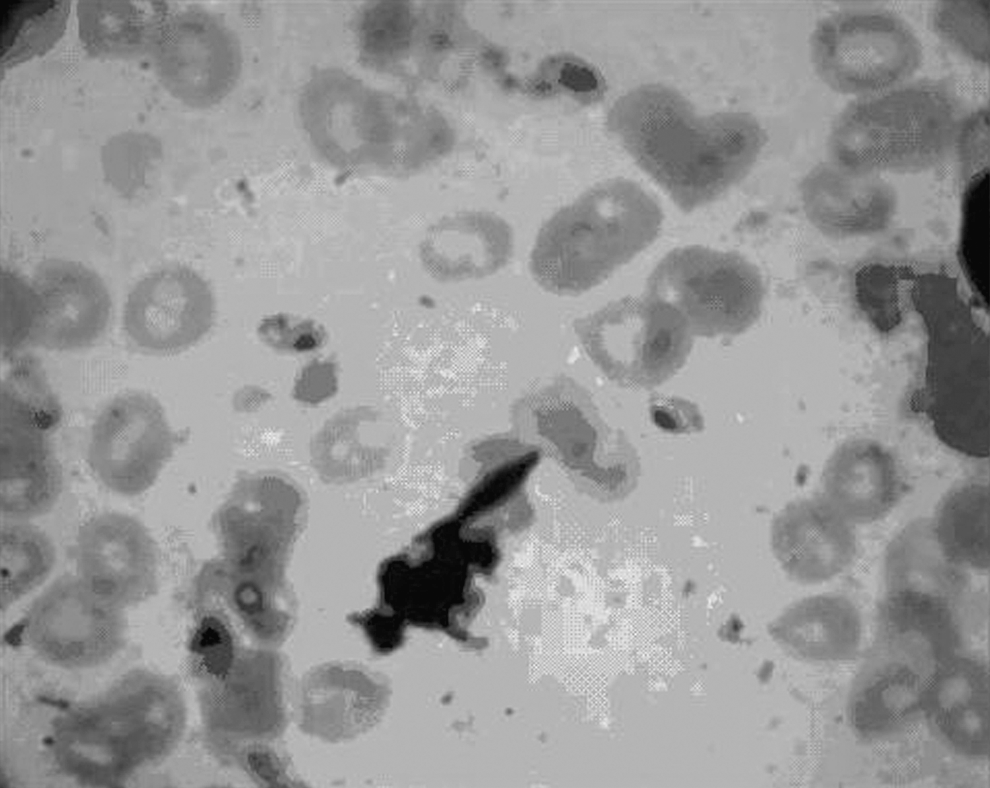
The amastigotes of Leishmania major from a gerbilline rodent.
Duplicate smears—the six found amastigote-positive and the four amastigote-negative by microscopy—were approved for Leishmania kDNA by PCR. No other slide smears were PCR-positive. All the detected kDNA appeared to come from L. major. For each PCR-positive sample, the second round products of the nested PCR were identical to those of the L. major reference strain, with a main band of 560 bp, and distinct from those of the L. infantum and L. tropica standards, which had main bands of 720 bp and 750 bp, respectively (Fig. 3).
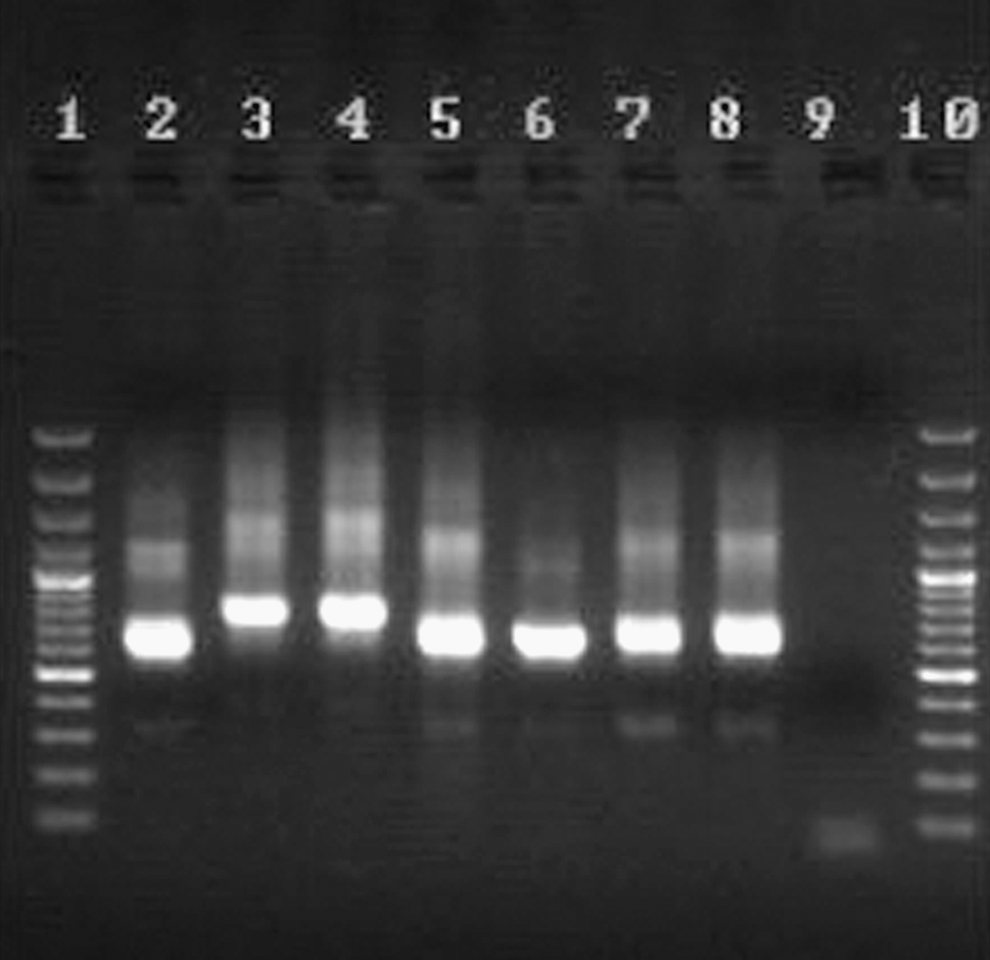
The results of the PCR-based amplification of kinetoplast DNA recovered from smear samples from Meriones hurrianae (lane 7), Tatera indica (lane 8), Gerbillus nanus (lanes 5 and 6), or from reference samples of Leishmania infantum (lane 4), L. tropica (lane 3), or L. major (lane 2). A negative control (lane 9), and molecular-weight markers (lanes 1 and 10) were also run to check for identity.
Discussion
Multiple numbers of host species are potentially involved in the transmission of Leishmania parasites, making its epidemiology highly complex (Emami et al. 2009). A major challenge facing those studying the epidemiology of the Leishmania species that cause zoonotic disease in humans is identification of the mammalian hosts that act as reservoirs of the involved pathogens. It is often difficult to collect large numbers of each putative wild rodent reservoir, or to successfully detect leishmanial infections because the parasites may cause no or only minor skin lesions in these hosts (Moemenbellah-Fard et al. 2003). Thus it is difficult to verify that the detected parasites are the ones that cause human disease (Pourmohammadi et al. 2008). It is, however, necessary to identify the parasite reservoir hosts, as they are often fundamental factors in the epidemiology and control of human disease (Oliveira et al. 2005).
Numerous species of rodents have thus far been reported from Iran. Some 70 species were caught by Edrissian and associates (1975), while 65 species of rodents were cited by Darvish (2005). In the present surveillance of the rodents' fauna, the trapped specimens were distributed in five wild (Meriones persicus, M. libycus, M. hurrianae, Tatera indica, and Gerbillus nanus) rodent species. No commensal rodent species were captured because sampling was carried out on the outskirts of the study villages.
Although <60% of the trapped rodents in this study were from the western plain, those from the eastern plain were more likely to be infected with Leishmania parasites. The latter ecotope lies next to the Sistan-Baluchistan province, with many foci of ZCL disease bordering Pakistan's littoral regions. Considering the province's pathogenic landscape topography and the patchy dispersion of infectious rodent reservoirs, it is likely that the infection spreads from east to west in this region. There are some preliminary unpublished data on the infection rate of P. papatasi in the eastern and western plains, which indicate a 5% positivity rate of infection with L. major using PCR in the eastern plain of the Jask region.
The fact that one species (M. hurrianae) was very common in the Jask region implicated its potential role as the main reservoir host of Leishmania parasites, similar to that found in zoonotic foci of the disease from the Rajasthan region in northwest India (Molyneux and Ashford 1983). M. hurrianae is the actual natural reservoir host of ZCL in the southeast region of Iran (Ashford 1996).
The eastern plain of the Jask region also had a relatively high (16%) abundance of free-living wild Baluchistan gerbils, G. nanus, a solitary nocturnal seed-eating desert rodent, the role of which in the eco-epidemiology of ZCL in Iran remains unclear. This was crucial, since no new records of it were available from this region in the last half century. It needs to be reassessed in the wake of more recent observations to help in planning control measures. The first report on the presence of G. nanus as a potential reservoir of L. major was recently published (Azizi et al. 2011), for a rural region that lies within its previously defined geographical limits in Iran, five decades after Misonne's report (1959). This finding is therefore of major epidemiological importance. In our study (Azizi et al. 2011), the first evidence of natural infection of G. nanus with the protozoan parasite L. major shed more light on the incrimination of putative zoonotic reservoirs of ZCL infection in this region. As G. nanus and M. hurrianae were both relatively common and positive for leishmanial amastigotes, it seems likely that they are the main rodent reservoir hosts in the study region, assuming that most of their leishmanial parasites belong to species causing the human ZCL disease observed in the region. The recent report of ZCL infection from a species of the Indian gerbil, T. indica, and two unspecified genera of Gerbillus s.l. in the southeast region of adjacent Fars province, should thus be reconsidered in the light of the results of the present study (Oryan et al. 2007).
Many molecular methods, in particular those based on PCR, have recently been used for the detection and identification of Leishmania species (Reithinger and Dujardin 2007; Selvapandiyan et al. 2008). The leishmanial DNA generally remains intact following the drying, fixation, and routine staining used to make tissue smears for microscopy (Motazedian et al. 2002). It is thus possible to use PCR to identify leishmanial species in fixed and dried Giemsa-stained smears (Laskay et al. 1995; Azizi et al. 2008). In this study, the scrapings from slide smears of rodent tissues subjected to PCR proved to be reasonably useful in revealing which Leishmania species were present. We concluded that M. hurrianae, G. nanus, and T. indica could act as potential reservoirs of L. major in the Jask district. The fact that no infection was observed in M. libycus and M. persicus does not preclude them from their known role as a reservoir of L. major.
Footnotes
Acknowledgments
The authors are grateful for the logistical support given by the Tropical and Infectious Diseases Research Centre, HUMS, Bandar Abbas, Iran. We appreciate the help with the field work done by Mr. S. Fekri, and for partial modification of ![]() by Ms. T. Dabagh. We are indebted to Ms. M. Adnafi for initial identification, and to Prof. J. Darvish for the expert verification of rodent identities. This article was the result of a research plan (no. M/P/251, dated 1-1-08), carried out under the auspices of HUMS. Finally, the authors are indebted to the vice-chancellor for research and technology at SUMS for permitting the use of facilities at this university. This study was conducted with the permission of the Natural Resources Authority, Environmental Protection Organization of Iran. All animal procedures were in accord with and approved by the Ethics Committee for the “Code of Practice on the Ethical Care and Use of Animals in Experiments” at HUMS.
by Ms. T. Dabagh. We are indebted to Ms. M. Adnafi for initial identification, and to Prof. J. Darvish for the expert verification of rodent identities. This article was the result of a research plan (no. M/P/251, dated 1-1-08), carried out under the auspices of HUMS. Finally, the authors are indebted to the vice-chancellor for research and technology at SUMS for permitting the use of facilities at this university. This study was conducted with the permission of the Natural Resources Authority, Environmental Protection Organization of Iran. All animal procedures were in accord with and approved by the Ethics Committee for the “Code of Practice on the Ethical Care and Use of Animals in Experiments” at HUMS.
Author Disclosure Statement
No competing financial interests exist.