
Abstract
We collected 284 ticks in Ethiopia (109 Amblyomma cohaerens, 173 Rhipicephalus decoloratus, and 2 Rhipicephalus praetextatus). We found no rickettsiae and bartonellae. In 7.3% of the A. cohaerens, we found a Borrelia sp. that may represent a new species distant from both relapsing fever group and Lyme borreliae.
Introduction
However, few studies have investigated the role of hard ticks as vectors of human pathogens in Ethiopia. The first evidence of the presence of the spotted fever group (SFG), rickettsia, and the agent of Q fever (Coxiella burnetii) in ticks collected from cattle in Ethiopia was reported in 1966. Several strains of SFG rickettsia have been isolated from Amblyomma variegatum and Rhipicephalus simus, and serological evidence of infection in cattle has also been found (Philip et al. 1966). One of these isolates, Ethiopian spotted fever rickettsia (ESF-5), was identified as Rickettsia africae (Philip et al. 1966; Kelly et al. 1996).
Among recent reports is a case of R. africae infection (African tick-bite fever) in a man who traveled to Ethiopia (Stephany et al. 2009). While in Ethiopia, he had assisted with a documentary film production about an Ethiopian tribe and had been in contact with cattle in the villages. Another epidemiological study reported the presence of R. africae in 12/118 Amblyomma lepidum ticks and in 1/2 A. variegatum ticks (Mura et al. 2008). Both species of ticks are classical reservoirs and vectors of R. africae (infection rates up to 100%) and are limited to low altitudes. Ticks of the subgenus Boophilus of the genus Rhipicephalus usually cohabit with A. variegatum and A. lepidum and are also often reported to be infected by R. africae, but at lower rates, usually at 10–20%.
A set of very interesting studies that have not yet been reproduced were published in the 1950s and 1960s. Ruth Reiss-Gutfreund reported on the isolation of the agent of epidemic typhus, Rickettsia prowazekii, from Amblyomma ticks in Ethiopia (Reiss-Gutfreund 1956, 1961, 1966). Although the isolated strains were identified as typhus rickettsia, follow-up efforts to repeat the isolation were not successful (Philip et al. 1966), and the hypothesis that Ethiopian cattle are reservoirs of typhus fever remains unconfirmed.
The present study on cattle ticks in the southwest Ethiopian highlands was aimed at determining their role as vectors of human pathogens by using molecular biology methods.
Materials and Methods
The study was performed in September, 2010, in 5 villages in Ethiopia (Table 1). The altitude varied from 1355 meters (Tum) to 2395 meters (Gibarku). Ticks were collected manually from the cattle and identified morphologically (Walker et al. 2003, Hoogstraal 1956). In total, we collected 284 ticks (Table 1), including 109 Amblyomma cohaerens, 173 Rhipicephalus (Boophilus) decoloratus, and 2 Rhipicephalus praetextatus. Both A. cohaerens and Rh. (B.) decoloratus were often found cohabiting on the same host.
DNA from homogenized ticks was extracted using a BioRobot MDx Workstation (Qiagen, Courtaboeuf, France) with a customized extraction protocol following the manufacturer's instructions. DNA was stored at 4°C until it was used in a PCR.
The DNA extracted from the ticks was tested by genus-specific quantitative real-time PCR (qPCR) for the presence of Rickettsia spp., Bartonella spp., and Borrelia spp. Rickettsial DNA was also screened by qPCR using a Rickettsia-specific gltA gene–based primers and probe (Rolain et al. 2002). All samples were also screened using a Bartonella-specific qPCR assay with a 21-bp probe that targeted the 16S/23S rDNA gene intergenic spacer (ITS) (Raoult et al. 2006). We used a 16S rDNA-based genus-specific qPCR for the detection of borreliae (Parola et al. 2011). The sequences of primers and probes were as follows: Bor_16S_3_F, 5′-AGCCTTTAAAGCTTCGCTTGTAG-3′, and Bor_16S_3_R, 5′-GCCTCCCGTAGGAGTCTGG-3′, for primers and Bor_16S_3_P, 6-FAM-CCGGCCTGAGAGGGTGAACGG-TAMRA, for the fluorescent probe.
Samples positive for borreliae in qPCR were subjected to a standard PCR in automated DNA thermal cyclers to amplify the portion of the flaB (flagellin) gene of Borrelia spp. (Vial et al. 2006) and borrelial noncoding intragenic spacer (IGS) (Bunikis et al. 2004). Water and DNA extracted from a pathogen-free laboratory culture of A. variegatum were used as a negative control in the reactions, and DNA from Rickettsia sibirica, Borrelia crocidurae, and Bartonella quintana were used as positive controls in the specific qPCR.
The amplified products were detected by electrophoresis on 2% agarose gels in Tris/borate/EDTA (TBE) 0.5× buffer and were stained with ethidium bromide. The remaining reaction mixtures were stored at −20°C for direct sequencing. Amplified DNA was sequenced using an ABI PRISM 3730xl DNA Analyzer (Genome Express, Grenoble, France).
The evolutionary history was inferred using a Bayesian phylogenetic analysis (Ronquist and Huelsenbeck 2003) by TOPALi 2.5 software (Biomathematics and Statistics Scotland) with an integrated MrBayes application (
Results
The screening of all ticks for Bartonella spp. and Rickettsia spp. produced no positive results. However, 8/109 (7.3%) of the A. cohaerens ticks produced positive results when tested for the presence of Borrelia spp. with a 16S-based genus-specific qPCR system. Four of the positive ticks were from Mizan Teferi collected from 2 cows, 2 were from Gibarku from the same cow, and 1 was from Tum and Tikemit Eshet. There were 3 males, 3 females, and 2 nymphs. No samples produced positive results when amplified with borrelial IGS primers. We succeeded, however, in the amplification of a 344-bp fragment of flaB gene from all 8 ticks that were positive in qPCR. A BLAST search showed that the sequence generated resembled Borrelia but had a very low identity level with all known strains, sharing only 85–86% with the following officially recognized species: Borrelia duttonii, Borrelia recurrentis, and B. crocidurae. It was deposited in GenBank under accession number JX089967. A phylogenetic analysis of a 297-bp portion of the flaB gene showed that this Borrelia forms a separate branch on the phylogenetic tree apart from the Lyme disease and recurrent fever groups (Fig. 1).
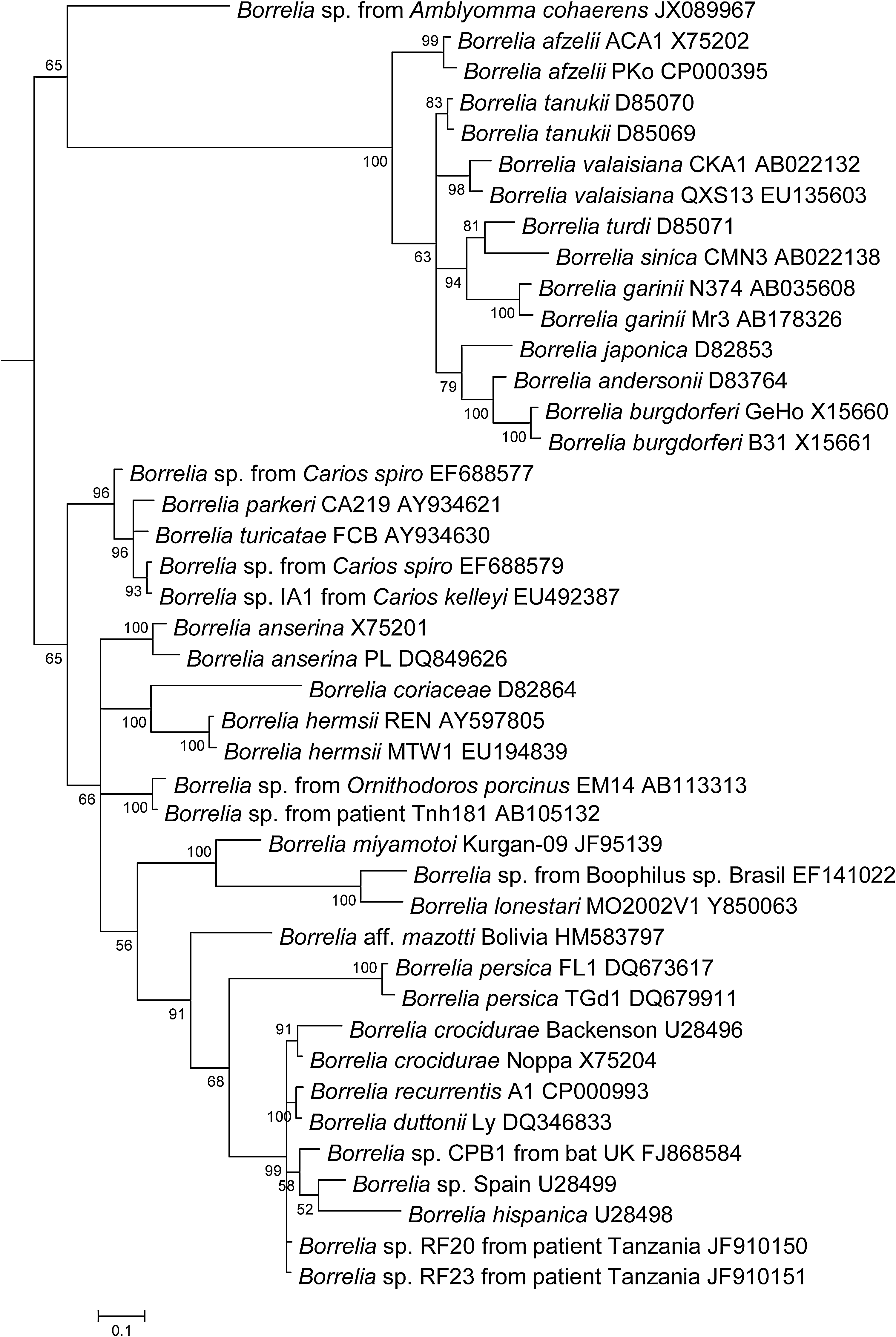
Phylogenetic relationships of 42 Borrelia. The evolutionary history was inferred using a Bayesian phylogenetic analysis with 1 substitution model for whole alignments based on flaB sequences. The confidence probability (multiplied by 100) that the interior branch length was greater than 0 was estimated using the bootstrap test (100 replicates is shown next to the branches). The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The strain, if available, is indicated after the species and followed by GenBank accession number.
Discussion
A. cohaerens has often been reported as the most abundant tick in cattle in southwestern Ethiopia, accounting for as much as 47.7% of all ticks collected (Abera et al. 2010). It has been recorded in areas between 1750 and 2500 meters above sea level that have an annual rainfall ranging between 750 and 1500 mm (Abera et al. 2010). This tick prefers high altitudes and moist climates and is absent in drier regions.
Rh (B.) decoloratus is not an altitude-dependent species and can be infected by R. africae when cohabiting with A. variegatum on the same bovine. We did not find R. africae in A. cohaerens ticks taken from cattle in the Ethiopian highlands. Several strains of R. africae were isolated from ticks (A. variegatum and Rh. simus) collected in southern Ethiopia in 1960s (Philip et al. 1966). However, despite of a large number of ticks tested by Philip et al. (72 ticks inoculated in animals), no strain was isolated from A. cohaerens and guinea pigs stayed afebrile; the only suggestion for the probable presence of rickettsiae was that the animals became immune to the following challenge. Similar to Philip et al., in our study, Rh. (B.) decoloratus was not infected by rickettsiae, although 173 were tested. This finding may suggest that Rh. (B.) decoloratus could be the secondary reservoir of R. africae, which becomes infected only when cohabiting with the primary reservoir A. variegatum, which is usually infected by R. africae in a very high degree, up to 100% (Parola et al. 2004; Mediannikov et al. 2010). In our study, in the higher altitudes, A. variegatum was replaced by A. cohaerens, which, apparently, did not carry R. africae. Consequently, no Rh. (B.) decoloratus infected by R. africae were identified.
We did not identify any Bartonella spp. in the ticks we studied. To the best of our knowledge, no data on Bartonella in hard ticks in Africa exist to date. However, different species, including these pathogenic for humans, were identified from lice (Angelakis et al. 2011), fleas (Rolain et al. 2005), soft ticks (unpublished data), ill humans (Mainardi et al. 1996), and animals (Chomel et al. 2006).
The Borrelia sp. identified in this study is of interest. A BLAST search of its flaB gene sequences showed that it differs significantly from the species found in East Africa. For instance, the identity with B. duttonii is only 86% and with Borrelia spp. recently identified in soft ticks Ornithodoros porcinus is only 84.5%. Both these borreliae are pathogenic for humans. The last one, “Borrelia mvumii” was isolated from the blood of acute febrile patients in Tanzania (Kisinza et al. 2003). Two other borreliae of veterinary importance were identified in ticks in Ethiopia. They are Borrelia anserina, avian pathogen, in Argas persicus soft ticks and Borrelia theileri, the cause of bovine borreliosis, a relapsing fever-like illness (Cutler et al. 2012). B. anserina was never identified in hard ticks; however, B. theileri is transmitted by the Rhipicephalus (Boophilus) hard ticks (Smith and Rogers 1998), but the genetically similar Borrelia lonestari is associated with Amblyomma spp. in North America.
The phylogenetic analysis (Fig. 1) of this bacterium shows that it formed a branch very distant from the clusters of two major groups of borreliae. The Borrelia sp. identified in A. cohaerens has an intermediate position between the recurrent fever group and the Lyme disease group. This new Borrelia sp. possibly belongs to a new third clade of the genus. Because many Borreliae are pathogenic in humans and animals, the potential role of this new Borrelia in human and animal pathology should be investigated.
Author Disclosure Statement
No competing financial interests exist.