
Abstract
The arrival of West Nile virus (WNV) in North America has led to interest in the interaction between birds, the amplification hosts of WNV, and Culex mosquitoes, the primary WNV vectors. American robins (Turdus migratorius) are particularly important amplification hosts of WNV, and because the vector Culex mosquitoes are primarily nocturnal and feed on roosting birds, robin communal roosting behavior may play an important role in the transmission ecology of WNV. Using data from 43 radio-tracked individuals, we determined spatial and temporal patterns of robin roosting behavior, and how these patterns related to the distribution of WNV-infected mosquitoes. Use of the communal roost and fidelity to foraging areas was highly variable both within and among individual robins, and differed markedly from patterns documented in a previous study of robin roosting. Although there were clear seasonal patterns to both robin roosting and WNV occurrence, there was no significant relationship between communal roosting by robins and temporal or spatial patterns of WNV-positive mosquitoes. Our results suggest that, although robins may be important as WNV hosts, communal roosts are not necessarily important for WNV amplification. Other factors, including the availability and distribution of high-quality mosquito habitat and favorable weather for mosquito reproduction, may influence the importance of robin roosts for local WNV amplification and transmission.
Introduction
For several reasons, American robins (Turdus migratorius) have been identified as a species important to the transmission of WNV. Robins experience low mortality from WNV and are viremic for 1–3 days (Kilpatrick et al. 2010). Analyses of mosquito blood meals (Apperson et al. 2002, 2004; Molaei et al. 2006; Savage et al. 2007; Patrican et al. 2007; Hamer et al. 2008, 2009), and field tests comparing mosquito host-preference for birds (Simpson et al. 2009), suggest robins are a preferred host. Robins may regulate the activity of WNV based on their abundance in an area and their migration or dispersal to new sites in the fall (Kilpatrick et al. 2006a, 2006b). Given this association between robins and WNV, the spatial ecology of robins is likely to influence WNV transmission. Because Culex mosquitoes are active throughout the night (Anderson et al. 2007), and encounter most bird species as they roost (Ward et al. 2006; Diuk-Wasser et al. 2010), the roosting behavior of robins may be particularly important in explaining spatial and temporal variation observed in the transmission of WNV. Our goal in this study was to use radio telemetry to document spatial and temporal patterns of roosting of American robins and their associated daytime movements, and to determine how those patterns relate to the spatial and temporal distribution of WNV-infected mosquitoes.
Like many bird species, individual robins regularly gather to spend the night together. Conventional theory suggests that benefits of such communal roosting (e.g., reduced predation and improved foraging; Lack 1968; Ward and Zahavi 1973; Weatherhead 1983) should cause birds to exhibit high fidelity to communal roosts. In a radio-telemetry study of American robins in New Jersey, however, exactly the opposite picture emerged (Morrison and Caccamise 1985, 1990). Individual birds were highly faithful to foraging areas, but changed communal roosts regularly. If robins generally behave this way, their potential to amplify and promote the spread of WNV would be much greater than if they maintained high fidelity to a communal roost. A robin infected with WNV would come in contact with many more other robins if it regularly switched communal roosts. Thus our first objective was to document the fidelity to a communal roost by robins in central Illinois, and to determine where they spend the night when not in the communal roost.
An important difference between our study and that of Morrison and Caccamise (1990) is that they tracked only adult robins, whereas we tracked both adults and juveniles. Inclusion of juveniles may be important for two reasons. First, juvenile robins are likely to play an important role in WNV transmission. Approximately 15% of adult robins in Illinois carry antibodies for WNV, indicative of having survived a previous infection and providing them immunity from future infection (Beveroth et al. 2006; R.L. Lampman, unpublished data), whereas all juveniles should be WNV-competent hosts. Second, the roosting behavior of juvenile robins is likely to differ from that of adults. Juvenile robins are less efficient at foraging than adults for months after juveniles have become independent (Gochfeld and Burger 1984; Vanderhoff and Eason 2007, 2008). Therefore, if robins use communal roosts to obtain foraging information (Ward and Zahavi 1973), juveniles should use communal roosts more than adults (Weatherhead 1983). Our second objective was to test that prediction.
Our third objective was to document the spatial and temporal pattern of mosquitoes infected with WNV, and relate that to the roosting behavior and movements of American robins studied in the same location. We first determine whether the proportion of mosquitoes infected with WNV increases with proximity to a communal roost. A recent study in Connecticut did find a higher proportion of infected mosquitoes at communal robin roosts than at non-roosting locations (Diuk-Wasser et al. 2010). We then examine the temporal patterns of mosquito infection. In Illinois, WNV activity is consistent from year to year, emerging in early summer and peaking in late August to early September (Huhn et al. 2005). If the number of robins using communal roosts affects seasonal patterns of WNV transmission, variations in the detection of WNV in mosquitoes should be correlated with variations in communal roosting by robins.
Materials and Methods
We conducted this study in Champaign, Urbana, and Savoy, three separate cities in Champaign County, Illinois, that functionally are a continuous urban area that encompasses the campus of the University of Illinois. The habitat was a mix of urban forest, residential lawns, second-growth forest, and agricultural fields associated with the university. The only large communal roost in the study area was in a mature cedar hedge that formed the western boundary of the university's pomology research area.
Radio tracking of robins
From July 2006 through October 2008, we radio-tagged 43 robins (30 adults, 9 juveniles, and 4 of unknown age). The robins were aged based on plumage (Sallabanks and James 1999). Because we only caught juveniles in late summer, and they either migrated or molted into adult plumage by late fall, data for juveniles were limited to July 27 to October 27. Robins were captured in second-growth forest, residential lawns, or suburban parks at 8 locations in Champaign (3), Urbana (4), and Savoy (1). Robins were captured in Savoy only in the second half of the study. We intentionally scattered capture locations to avoid concentrating captures in a particular location or habitat. All robins were captured by passive mist nesting (i.e., no effort was made to attract birds to the nets). We generally limited tracking to 4 birds at a time, and once a transmitter failed or fell off an individual, we selected a new area and individual. No individual robin was radio-tagged more than once in the study. Because migration reduces the size of the local robin population late in the fall, we tracked fewer birds late in the season.
We attached a 1.5-g radio transmitter to the central rectrices of each bird using heat shrink tubing (Alessi et al. 2009). We attempted to locate all birds with transmitters every day and night, although holidays, snowstorms, and vehicle breakdowns resulted in a small number of missed tracking days. For robins that retained transmitters and did not migrate, we located individuals on >95% of nights. At night the birds were located ≥1 h after sunset. Daytime locations were alternated daily between morning (08:00–10:00 h) and afternoon (13:00–15:00 h), and were occasionally determined for both periods. We used two approaches to locate birds. First, we used an automated radio telemetry system to guide us to the general location of a tagged bird. This approach also allowed us to determine if an individual migrated from the area (Ward and Raim 2011). Second, we used a directional antenna mounted on a vehicle to determine specific locations by recording several directions and triangulating. Positions in the field were determined using a 2- to 3-m resolution GPS unit mounted in the vehicle. Ground truthing of approximately 30 roost and 50 daytime locations by hand-held radio tracking indicated that locations estimated by triangulation were within 25 m of the actual locations.
Sampling mosquitoes and WNV detection
We sampled mosquitoes using Centers for Disease Control and Prevention (CDC) gravid traps infused with grass, which are designed to attract gravid Culex females (Lampman et al. 2006). We placed 23 traps each year from 2003–2008 in a variety of habitats throughout the study area, including residential, park, commercial, campus, livestock, and woodlots. Trapping started in May or June of each year, and ended in September or early October. The traps were checked every week. Mosquitoes were stored in 2.0-mL Eppendorf tubes in batches ranging from 1–50 mosquitoes at −80°C until analyzed for the presence of WNV. Each batch contained mosquitoes from only a single sampling location and week.
We used standard methods (Lampman et al. 2006) to detect WNV in batches of mosquitoes. Briefly, reverse-transcription polymerase chain reaction (PCR) was used to detect the presence of WNV-RNA. RNA is first separated from samples, purified, and then subjected to real-time PCR amplification. Positive RNA controls from a WNV strain (NY99), and local WNV-positive mosquitoes, as well as 8 negative controls (no flavivirus RNA extract was added), were added to each plate to ensure accuracy.
Statistical methods
A bird was classified as roosting communally if it spent the night in the cedar hedge that comprised the only known communal roost in the study area. To account for potential GPS error, we used an aerial photograph to digitize the cedar-hedge boundaries, and classified individuals roosting within 25 m of the boundaries as attending the communal roost. No other large communal roosts were found, although some birds roosting elsewhere may have roosted with a small number of conspecifics. Of the 13 observations of robin departures from roosts other than the large communal roost, 11 were alone, and 3 were with 2 or 3 conspecifics. When we had roost and subsequent day-use locations, we calculated the distance between these points as a measure of the minimum distance moved. When data were available, we calculated the distance between locations determined on consecutive days. For all birds with ≥15 day-use locations, we estimated 95% fixed-kernel home ranges. Smoothing parameters were calculated using least-squares cross-validation (Kernohan et al. 2001).
We examined distances between roost and subsequent day-use locations and consecutive day-use locations using repeated-measures analysis of variance (Littell et al. 2006). We treated the individual bird and roosting status (communal or solitary) within bird as random effects, and roosting status, season, and their interaction as fixed effects. Seasons included pre-breeding (February and March), breeding (April through July), and two post-breeding periods (August and September, and October through December). We followed tests with significant main or interaction effects with pair-wise contrasts (Littell et al. 2006). Data were log-transformed to fulfill the normality and homogeneity of variance assumptions, but we present untransformed values for ease of interpretation.
To examine seasonal changes in communal roosting, we calculated the proportion of nights at the communal roost and the median tracking date for each individual. We examined effects of median tracking date, age, and their interaction, on the proportion of nights spent at the communal roost using logistic regression (Allison 1999). For the period when both adults and juveniles were tracked (July 27 to October 27), we examined differences in the proportion of individuals of each age-class that used the communal roost using a chi-square test. Similarly, for the same time period we used a generalized linear mixed model (Littell et al. 2006), to examine the daily probability of an individual using the communal roost as a function of age, day of the year, and their interaction, using the binomial distribution and logit link function. Again, we treated individual as a random effect, and used pair-wise contrasts to explore main and interaction effects (Littell et al. 2006).
To determine if communal-roost use affected home-range size we used analysis of covariance (Littell et al. 2002) to examine differences in home-range size (log-transformed) between birds that used communal roosts <50% versus ≥50% of nights, with median tracking date as a covariate to account for potential seasonal changes in home-range size.
For examining the relationship between WNV prevalence and spatial or temporal patterns of communal roost use by robins, we used mosquito data collected beginning in 2003. Even though we began tracking robins in 2006, we used previously collected mosquito data because long-term roost use is common for robins (Speirs 1946; James 1957), and the communal roost we studied had been used for ≥10 years before our study (M.P. Ward, personal observation). We examined relationships between WNV prevalence in mosquito traps, and both the distance from the communal roost, and the proportion of the robin population using the communal roost, during a bi-weekly interval using linear regression (Littell et al. 2002).
Results
We recorded locations of 43 radio-tagged robins for an average of 33.1 days, although we tracked some individuals for up to 135 days. Variation in tracking periods resulted from 11 birds migrating, and from premature transmitter failure or loss. We located robins on about 90% of attempts. Failure to locate birds probably resulted from them foraging or roosting near buildings that blocked signals. We collected 903 day-use and 1136 nocturnal locations between July 8, 2006 and December 11, 2008. We were able to calculate the minimum distance between a roost and subsequent day-use location on 623 bird-days, and measured 537 minimum distances between areas used on consecutive days (sample sizes by age and roosting status are summarized in Table 1).
The range is the minimum and maximum number of locations for that individual.
SE, standard error.
Qualitatively, the most apt description of robin roosting behavior is that it is highly variable, both within and among individuals. Their roosting behavior was not consistent with either conventional theory (high communal-roost fidelity, and low fidelity to day-use areas), nor did they behave similarly to the robins studied by Morrison and Caccamise (1990) in New Jersey (low roost fidelity and high fidelity to day-use areas). For individuals with ≥15 roosting observations, use of the communal roost ranged from 0–90% (mean 31%) of nights. For individuals with ≥15 daytime observations, home-range size varied from 1.3–14,417.1 ha (mean 1173.6 ha). We illustrate this variation with maps of day-use and roosting locations for two adult males with very different behaviors (Fig. 1). One male roosted almost exclusively in the communal roost and foraged over a wide area, whereas the other roosted in numerous locations, not including the communal roost, and restricted foraging to an area an order of magnitude smaller than the area used by the first male. Most individuals exhibited combinations of roosting and foraging between these extremes. Despite this variation in individual behavior, some patterns did emerge.
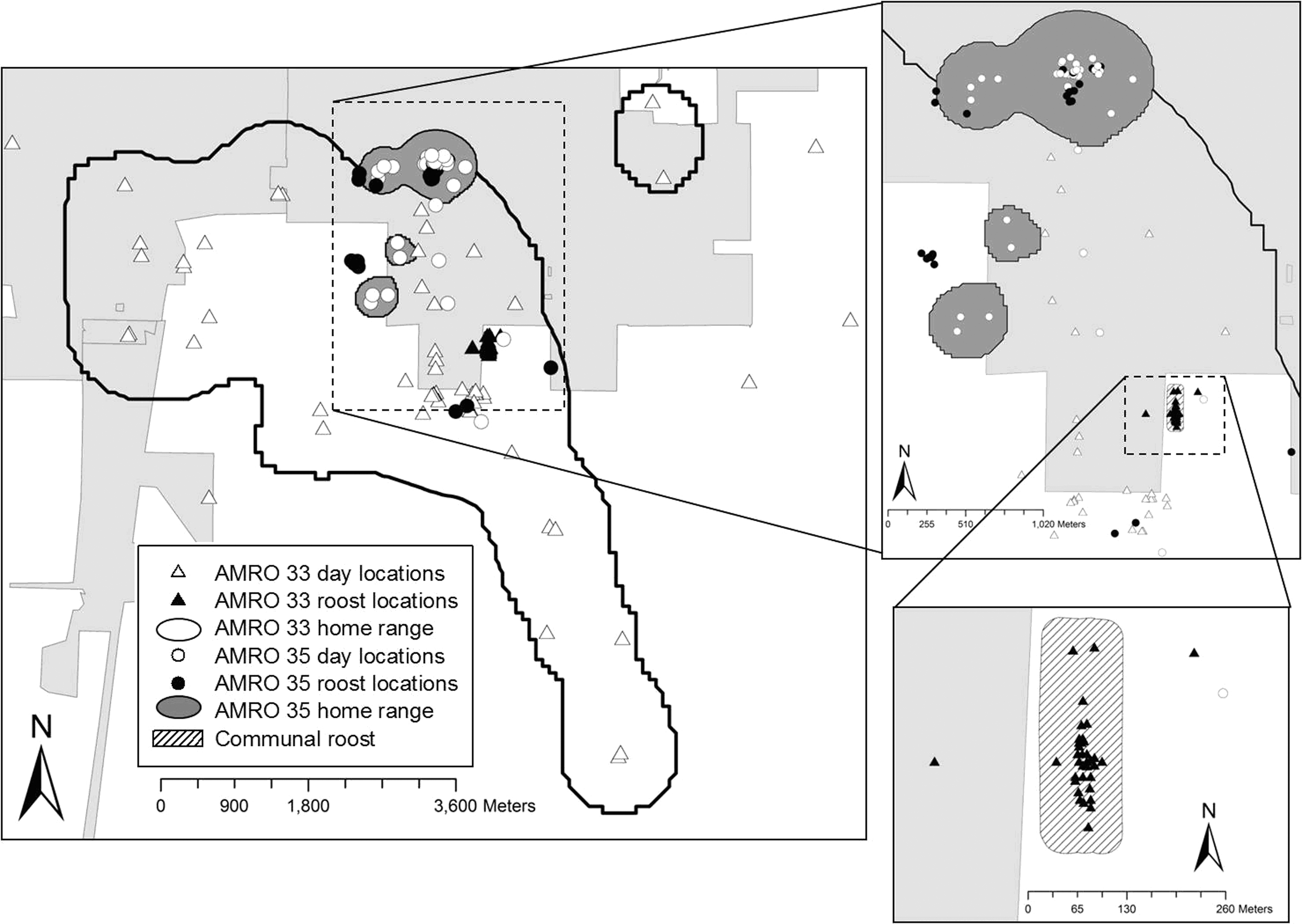
Contrasting space use and roosting patterns of two adult male American robins (AMRO 33 and 35) in the adjoining cities of Champaign, Savoy, and Urbana, Illinois (light gray shading), 2006–2008. Home ranges are 95% fixed-kernel estimates based on day-use locations. Sample sizes were 58 day and 40 roost locations, and 41 day and 32 roost locations, for AMRO 33 and 35, respectively. AMRO 33 was tracked in October and November of 2008, and AMRO 35 was tracked in June and July of 2008.
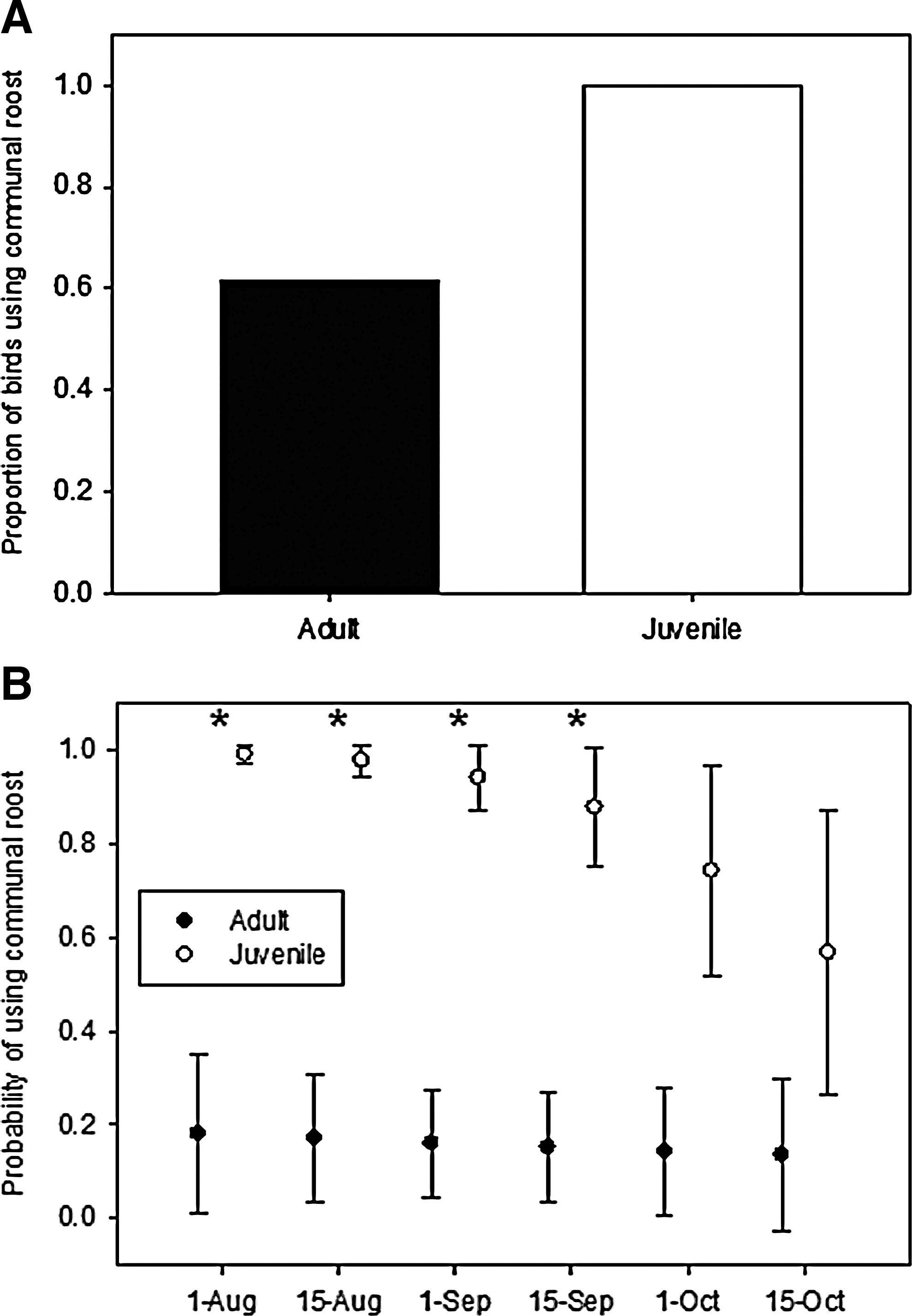
The proportion of roosting locations that were at the communal roost increased through the year, reaching a maximum in the fall (chi-square=151.97, p<0.0001; Fig. 2). During the time that both adults and juveniles were tracked (late July to late October), a smaller proportion of adults (62%) than juveniles (100%) used the communal roost (chi-square=4.04, p=0.04; Fig. 3A), and the proportion of nights at the communal roost differed between age classes (chi-square=44.76, p<0.0001). There was a weak interaction between day of year and age class in the probability that an individual used the communal roost on a given night from late July to late October [F(1,44)=2.81, p=0.10]; communal-roost use was high for juveniles for this entire period, but decreased slightly through the fall, whereas the probability for adults remained consistently low (Fig. 3B).
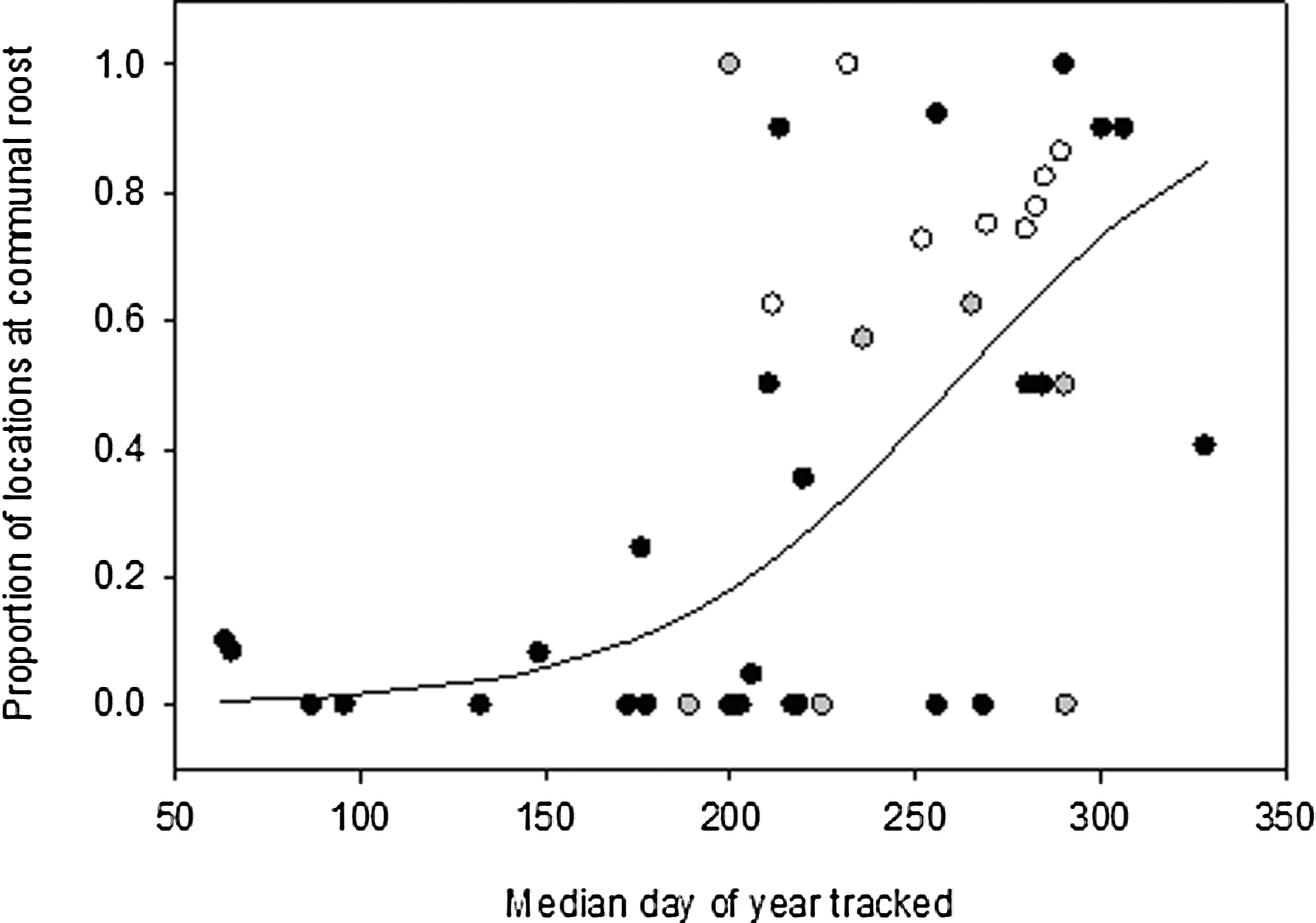
Proportion of American robin roosting locations that were at the communal roost as a function of the median date that an individual was radio-tracked for adults (black), juveniles (white), and individuals of unknown age (gray). Median date was significantly associated with the proportion of communal-roost locations (chi-square=151.97, p<0.0001). The equation for the fitted line is: ln[p/(1 − p)]=0.028×day − 7.12.
(
On consecutive days communal roosting robins foraged farther apart than when they roosted alone, and the distance between foraging areas used on consecutive days also increased over the year [F(1,21)=3.92, p=0.06, F(3,115)=10.32, p<0.0001, respectively; Fig. 4]. These observations are consistent with robins using the communal roost to locate new foraging areas, and the association was also apparent in the increase in day-use home ranges. We calculated day-use home ranges for 22 birds based on sample sizes that ranged from 16 to 85 observations (mean 36.5, SE 4.2). There was no relationship between sample size or sampling duration and home-range size [F(1,20)=0.05, p=0.83 and F(1,20)=2.46, p=0.13, respectively]. Day-use home ranges were bigger for robins that roosted communally ≥50% of the time than for those that used the communal roost <50% of the time [F(1,20)=14.18, p=0.0012]. Because communal roost use and tracking date were correlated, these variables could not be used together in the same model to predict home-range size. When included separately, however, communal-roost use and median tracking date had comparable r 2 values (0.41 and 0.43, respectively).
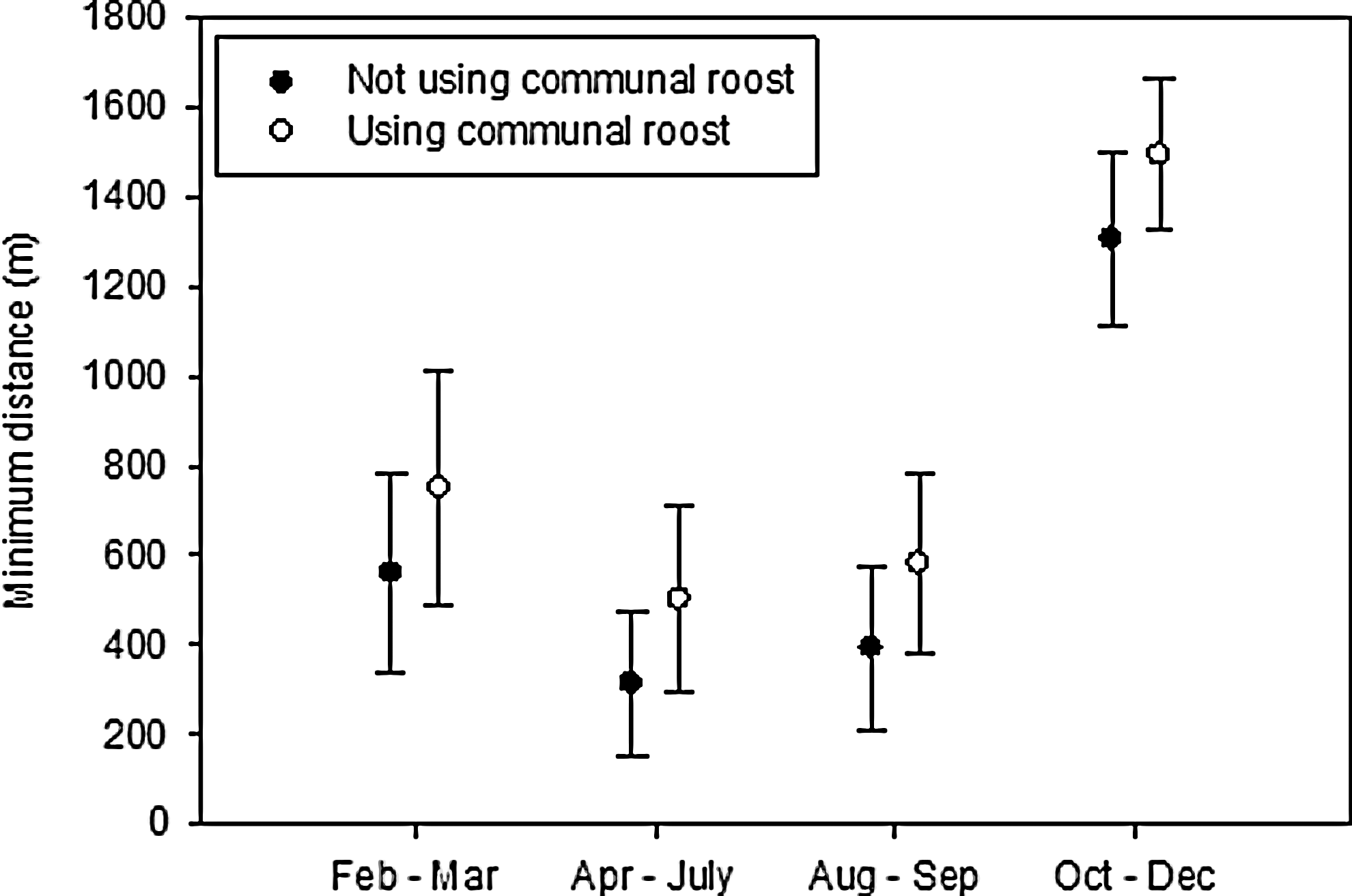
Minimum distance (m) between day-use areas attended on consecutive days for individuals that used the communal roost and those that roosted alone. The error bars indicate standard error.
Out of approximately 2900 pooled mosquito samples from 23 sites sampled consistently from May to early October 2003 to 2008, there were 135 positive samples. There were no WNV-positive samples in 2008, so we focused on data from 2003–2007 for temporal and spatial analyses. We found a clear seasonal pattern of the occurrence of WNV-positive mosquitoes: WNV was first detected in June, peaked in August, and remained common in September (Fig. 5). Despite year-to-year variations, the seasonal pattern was similar among years, and because we were interested in a stable robin roost, we focused on averaged data. WNV prevalence in mosquitoes varied among locations, but we detected WNV throughout the area sampled (Fig. 6). That broad distribution resulted in no association between WNV prevalence and distance from the communal roost (r 2=0.0001, p=0.96, Fig. 7). There was also no significant distance by year interaction (p=0.55). Temporally, WNV prevalence tended to increase with the bi-weekly proportion of robins using the communal roost, but not significantly (r 2=0.21, p=0.16).
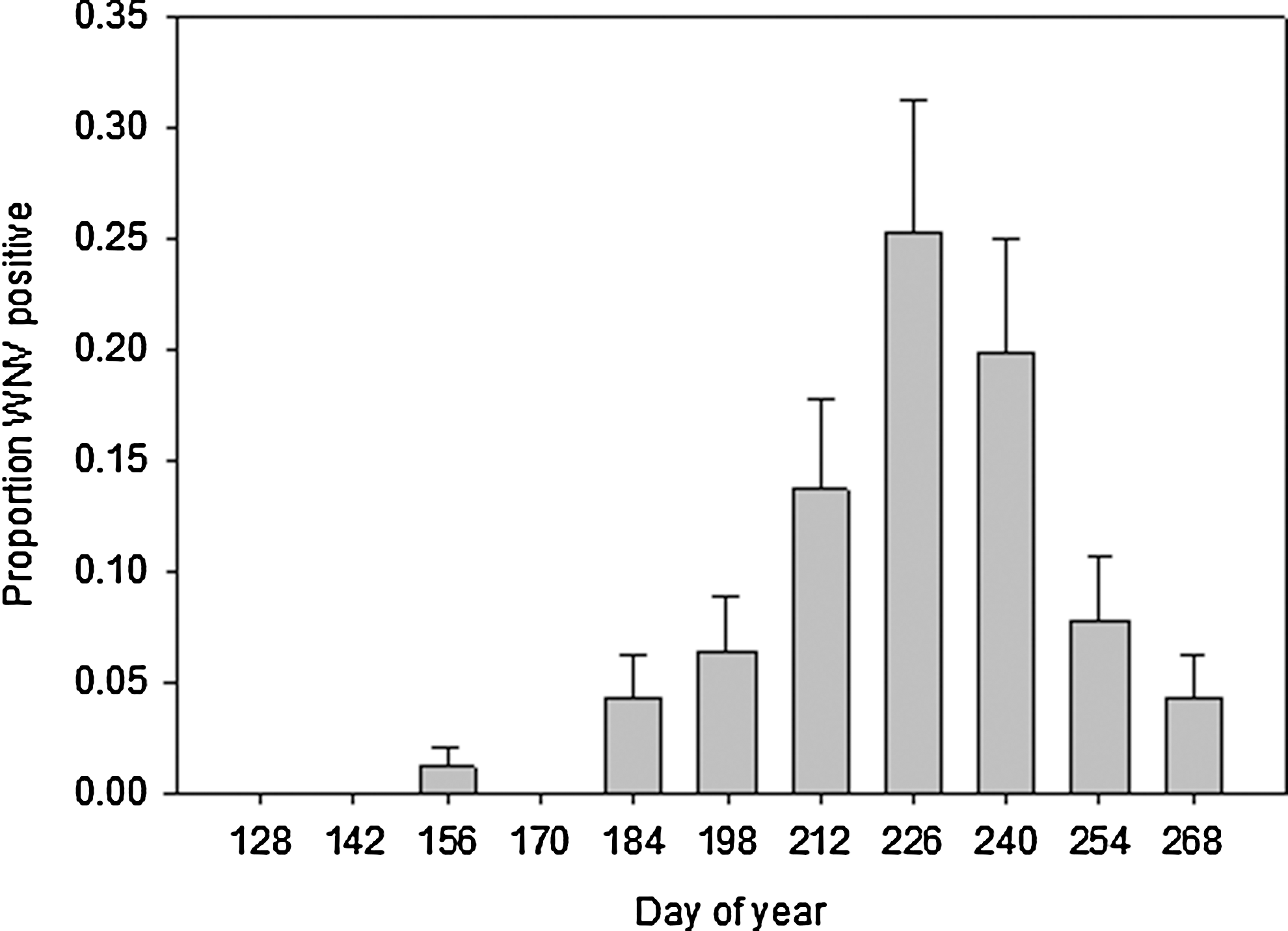
Proportion of samples with West Nile virus (WNV)-positive mosquitoes at 2-week intervals. The data are based on weekly samples from traps in place from mid-May until the first week of October, 2003–2007.
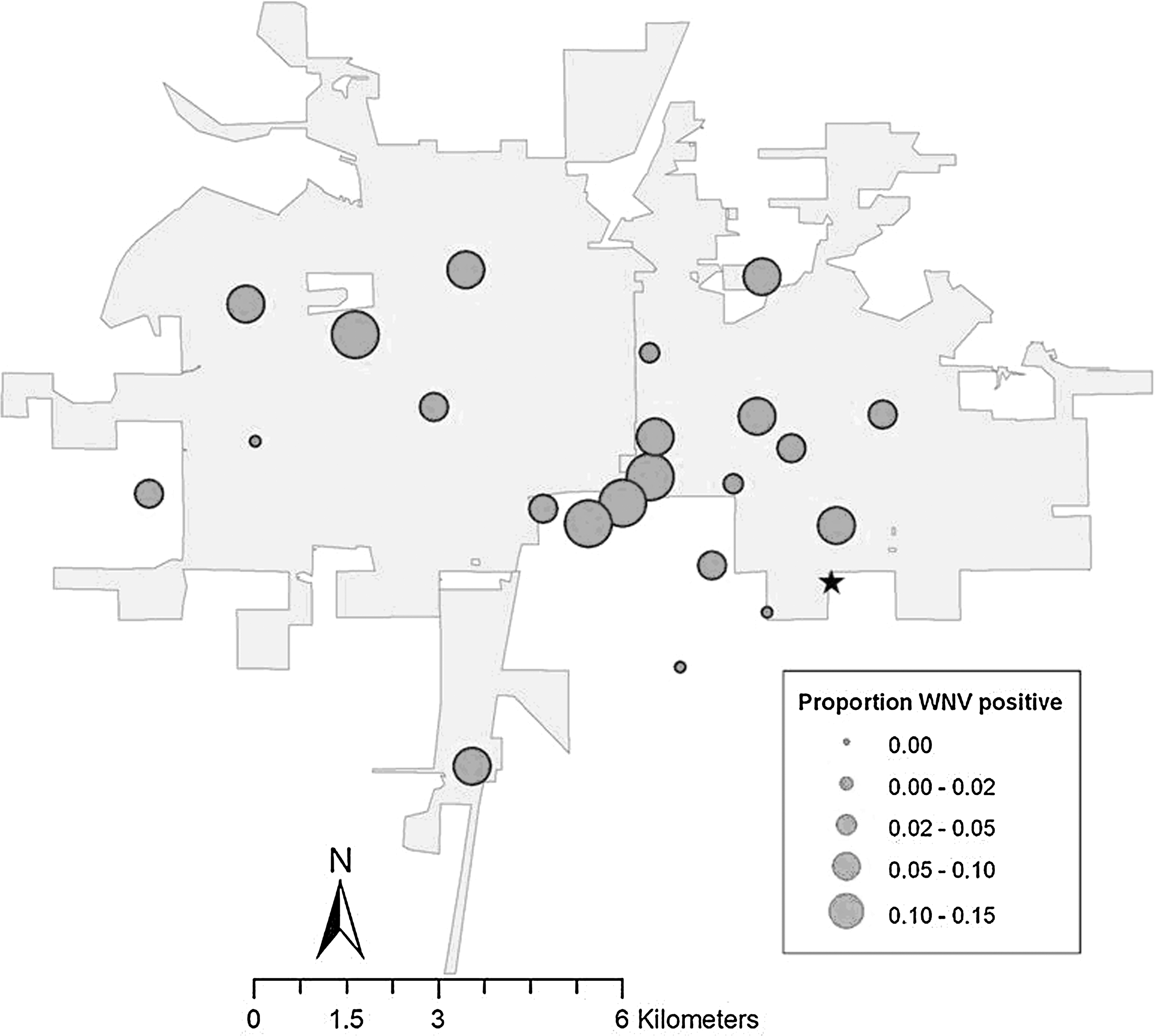
Proportion of weekly samples with West Nile virus (WNV)-positive mosquitoes across sampling locations in Champaign-Urbana (shaded area). The communal roost is designated by the star.

Proportion of samples with West Nile virus (WNV)-positive mosquitoes based on distance of the sampling location from the communal roost. The data are based on traps checked weekly from mid-May until the first week of October, 2003–2007.
Discussion
Roosting behavior of American robins in Illinois did not conform to either the expectation based on theory (i.e., high roost fidelity and low fidelity to foraging areas; Ward and Zahavi 1973; Weatherhead 1983), or to patterns reported for robins in New Jersey (i.e., low roost fidelity and high fidelity to foraging areas; Morrison and Caccamise 1990). Rather, roosting behavior varied substantially among individuals, with some robins roosting communally on 90% of nights and others always roosting solitarily. Similarly, fidelity to foraging areas ranged from low to high, and the area of daily foraging areas varied by more than an order of magnitude. Despite this variation, there was a clear increase in communal roosting over the summer and into the fall, and greater use of the communal roost by juvenile robins than by adults. Given the seasonal increase in WNV prevalence in Culex mosquitos, both the seasonal increase in communal roosting and the high rate of communal roosting by juvenile robins should have promoted an association between the communal roost and WNV prevalence in mosquitos. However, we found no evidence of such an association. WNV prevalence did not vary significantly with distance from the communal roost or with temporal variations in robin roosting behavior.
We observed a number of patterns that are consistent with robins using communal roosts to locate food (more communal roosting by juveniles, more communal roosting later in the season, foraging areas used on consecutive days were farther apart after robins roosted communally, and home ranges were bigger for robins that used the communal roost more). From the perspective of how roosting behavior might influence WNV transmission, however, the use of the communal roost provided the potential for the roost to affect WNV transmission. The important issue, therefore, is to understand why that potential was not realized.
One reason that we did not find the same association between WNV and communal roosting by robins reported by Diuk-Wasser and associates (2010) could be that the Culex-robin-WNV relationship in Illinois is different from that of other places robins have been studied. However, the importance of robins for WNV transmission documented from blood meal analyses of vectors, serology of avian species, and attraction of mosquitoes to caged live birds found elsewhere (Apperson et al. 2002, 2004; Molaei et al. 2006; Savage et al. 2007; Patrican et al. 2007; Simpson et al. 2009), has also been found in Illinois (Hamer et al. 2008, 2009; R.L. Lampman, unpublished data). Even given that the Culex-robin-WNV relationship appears to be relatively constant geographically, the relative importance of robins for WNV transmission in a particular location will depend on the abundance of robins and other competent hosts, so it is possible that differences in the avian communities in Illinois and Connecticut contributed to the difference in the results seen between the Diuk-Wasser group (2010) and our group. There is also the potential that the type of trap used to capture mosquitoes could influence the type (infected or not infected) of mosquitoes captured. In the Diuk-Wasser study (2010), they used both CO2 and gravid traps (results were not reported separately by trap type), whereas we used only gravid traps. There are two reasons that differences in the traps used are unlikely to explain the different results. First, CO2 traps have a much lower probability of capturing WNV-infected mosquitoes (Williams and Gingrich 2007), so it seems likely that a substantial number of WNV-positive mosquitoes in the Diuk-Wasser study may have come from gravid traps. Second, in order for the use of CO2 traps to explain the differences seen between studies, the CO2 traps would have had to capture predominantly local mosquitoes, whereas mosquitoes captured in gravid traps would have to have traveled long distances. Assuming that being gravid constrains flight of female mosquitoes more than not being gravid, greater movement by gravid females seems unlikely. In the case of our results, for gravid females to have fed at the roost and then traveled far enough from the roost before being captured to obscure any relationship between WNV and distance from the roost traveled to change the results of the correlation between distance to the roost and the proportion of WNV-positive mosquitoes (i.e., Fig. 7), would require that gravid females would have had to become infected at the roost, and then traveled thousands of meters in random directions, which again seems highly unlikely.
A potentially more promising explanation for the difference between studies may be that the association between WNV and communal roosts could depend on whether a communal roost is located in a good mosquito habitat. Culex pipiens generally oviposits in urban containers with high organic content (Lampman and Novak 1996; Trawinski and Mackay 2010). The communal roost in our study was located on a small, well-drained hill. This meant that there were probably few oviposition sites for mosquitoes near the roost, consistent with the traps near the roost capturing few Culex mosquitoes. In the study by Diuk-Wasser and colleagues (2010) in Connecticut, only one of seven communal roosts had high WNV activity, while five roosts had WNV activity similar to control locations. Thus, the association between roosting robins and WNV in that study was largely a consequence of a single roost. Whether that roost was in a better mosquito habitat relative to the other roosts was not reported. If the potential for communal robin roosts to amplify WNV depends on roosts coinciding with high-quality mosquito habitats, it is also likely that the amplification potential for a given roost could also vary annually, depending on rainfall. During wet years, mosquito oviposition sites may be more plentiful, increasing the WNV amplification potential of roosts that have few mosquitoes in dry years. In hot, dry summers, however, the importance of a roost being located in a good mosquito habitat could be very high, because hot and dry conditions may be ideal for both communal roosting and WNV activity. In dry years, soil invertebrates would be difficult for robins to obtain (Gochfeld and Burger 1984), and they may rely more heavily on fruit, a sporadically-distributed resource that would likely increase the benefit of roosting communally. In dry years mosquitoes may be more concentrated in permanent standing water (i.e., a good mosquito habitat). During these dry periods, catch basins that are rich in organic material often serve as the main breeding sites for urban Culex mosquitoes. High temperatures during these periods also shortens the extrinsic incubation period in mosquitoes, potentially increasing transmission in an area (Petersen and Roehrig 2001).
Our results, together with results from other studies, suggest that the established importance of robins as WNV amplifiers does not mean that all robin communal roosts are always important for WNV amplification. Communal roosts in good mosquito habitats under the correct weather conditions may result in areas of significant WNV amplification, but not all roosts will be associated with high WNV amplification. Communal roosts may vary in importance with the fidelity of robins to the roost, which in turn may be a function of the availability and distribution of the food on which robins feed. Because juvenile robins show greater communal roost fidelity than adults, juveniles may potentially be more important than adults as WNV amplifiers. Future research that addresses both the movements of robins and Culex mosquitoes will further improve our understanding of WNV transmission ecology.
Footnotes
Acknowledgments
We thank T.A. Beveroth and M.N. Barron for capturing birds and attaching transmitters. Radio tagging of American robins was conducted under the Institutional Animal Care and Use Committee protocol no. 06239.
Author Disclosure Statement
This work was funded by the Illinois Waste Tire Act Fund.