
Abstract
Chimeric yellow fever 17D/DENV-1-4 viruses (CYD-1–4) have been developed as a tetravalent dengue vaccine candidate which is currently being evaluated in efficacy trials in Asia and America. While YF 17D and DENV are mosquito-borne flaviviruses, it has been shown that CYD-1–4 do not replicate after oral infection in mosquitoes and are not transmitted to new hosts. To further document the risk of environmental dissemination of these viruses, we evaluated the replication of CYD-1–4 in ticks, the vector of tick-borne encephalitis virus (TBEV), another member of the flavivirus family. Females of two hard tick species, Ixodes ricinus and Rhipicephalus appendiculatus, were inoculated intracoelomically with CYD-1–4 viruses and parent viruses (DENV-1–4 and YF 17D). Virus persistence and replication was assessed 2, 16, and 44 days post-inoculation by plaque titration and qRT-PCR. CYD-1–4 viruses were detected in I. ricinus ticks at early time points post-inoculation, but with infectious titers at least 100-fold lower than those observed in TBEV-infected ticks. Unlike TBEV, complete viral clearance occurred by day 44 in most ticks except for CYD-2, which had a tendency to decline. In addition, while about 70% of TBEV-infected I. ricinus nymphs acquired infection by co-feeding with infected tick females on non-viremic hosts, no co-feeding transmission of CYD-2 virus was detected. Based on these results, we conclude that the risk of dissemination of the candidate vaccine viruses by tick bite is highly unlikely.
Introduction
Ticks comprise two major families, soft ticks (Argasidae), and hard ticks (Ixodidae), both of which can transmit or be reservoirs of flaviviruses (Labuda and Nuttall 2004). Unlike mosquitoes, ticks are characterized by a long life cycle of several months to over 2 years. To accomplish their life cycle, ticks need blood meals from a vertebrate host at each stage (larva, nymph, and adult). Transstadial and vertical (transovarial) transmission of flaviviruses within tick populations has been demonstrated, and events during tick molting affect virus survival and replication (Nuttall and Labuda 2003). Non-viremic transmission—the transmission of a virus from infected to non-infected ticks co-feeding on the same vertebrate hosts without detectable viremia—has been demonstrated and represents another important mechanism of virus circulation (Labuda et al. 1993a, 1993b; Nuttall and Labuda 2003). Under natural conditions, ticks become infected by feeding on an infected host, or by co-feeding in aggregations with infected ticks on the same host. In laboratory conditions, ticks, unlike mosquitoes, will not feed via blood tubes, and are routinely infected via intracoelomic inoculation. Although this method bypasses the natural gut barrier that represents an important selective force after oral infection, it has been demonstrated that inoculated TBEV penetrates tick salivary glands and replicates to significant levels within a few days, allowing ticks to efficiently infect hosts and co-feeding ticks (Labuda et al. 1993a, 1993b).
Dengue infection is caused by four serologically distinct DEN virus serotypes (DENV-1–4) transmitted by Aedes mosquitoes, and represents a growing health problem (Guzman et al. 2010). No dengue vaccine is licensed yet, but several candidates are currently under clinical evaluation (Coller and Clements 2011). Among them, the most advanced is the CYD tetravalent dengue vaccine, which is currently being evaluated in efficacy trials in humans (Guy et al. 2011). The tetravalent CYD dengue vaccine is composed of four live, attenuated, monovalent chimeric vaccine viruses, CYD-1–4, constructed by replacing the genes encoding YF 17D virus (vaccine strain) premembrane (prM) and envelope (E) proteins, with those of the four DENV serotypes. The CYD viruses thus possess a DENV envelope responsible for their tropism, together with the YF17D replication machinery.
The ability of CYD viruses to replicate in Aedes albopictus mosquito cell culture (C6/36), and in A. aegypti and A. albopictus mosquitoes, was previously evaluated in comparison with the parental YF 17D and wt dengue viruses. Virus replication was studied in mosquito bodies versus heads to assess dissemination. The CYD viruses were able to replicate in Aedes bodies after intra-thoracic inoculation, albeit at lower titers than wt dengue viruses, as seen with YF 17D, and only poorly infected mosquitoes after an infectious blood meal (Johnson et al. 2002, 2004; Higgs et al. 2006).
After mosquitoes, ticks have the highest potential to transmit flaviviruses. The present study was therefore designed to evaluate replication and transmission of CYD-1-4 viruses in these arthropods. Two different hard tick species were selected to represent the existing variability within the Ixodidae family: Ixodes ricinus (Prostriata), a Palaearctic species and the principal vector of the European TBEV subtype, and Rhipicephalus appendiculatus (Metastriata), a tick species found in Southeastern Africa. This latter tick species has no known vector competence for flaviviruses in nature, but is able to acquire TBEV following intracoelomic inoculation, and transmit it to uninfected co-feeding ticks (Labuda et al. 1993a).
Materials and Methods
Cells and viruses
African green monkey kidney cells (Vero76) were grown in IMDM (Gibco, Carlsbad, CA)/4% fetal bovine serum (FBS) at 37°C. Porcine kidney cells (PS) were grown in M199 (Gibco)/7% FBS at 37°C. The Czech TBEV strain Hypr (accession number U39292) was propagated through mouse brain passages. YF 17D attenuated virus (YF 17D-204 strain), DENV-1–4, and CYD-1–4, were propagated on Vero76 cells.
Virus titration
The YF 17D, DENV-1–4, and CYD-1–4 viruses were titrated on Vero cells in a standard immuno-plaque assay. Infectious foci were visualized after incubation with the pan-flavivirus mAb4G2 (Gentry et al. 1982). TBEV was titrated on PS cells, on which it forms lysis plaques that are easily detectable by staining with crystal violet.
Laboratory animals
Adult outbred ICR mice (weight ∼20 g), and adult outbred guinea pigs (weight ∼200 g) were purchased from Dobra Voda Breeding Station, Institute of Experimental Pharmacology and Toxicology, Slovak Academy of Sciences (SAS). Adult (15–20 g) BALB/C mice were purchased from Masaryk University Brno, Czech Republic. The animals were housed at the Institute of Virology, SAS, under conditions (cages of a given size, and food and water ad libitum) compliant with the applicable regulations (Act of the Government of the Slovak Republic from July 9, 2003 regulating the use of experimental animals), in an approved experimental facility (no. 12284/03-220). Euthanasia of the animals at the end of the experiments was done under deep anesthesia using diethyl-ether, by cervical dislocation.
Ticks colonies
Colonies of ticks have been maintained at the Institute of Zoology (IZ SAS) laboratory and kept in desiccators at room temperature with relative air humidity of 85%; field-collected ticks have been continuously added to improve the quality of the I. ricinus colony. Only the F1 generation of laboratory-bred I. ricinus females was used for virus inoculation. The colony of R. appendiculatus was initially established with ticks provided by the National Environment Research Council (NERC) Centre for Ecology & Hydrology (CEH), Oxford, U.K. For routine colony maintenance, the ticks were fed on laboratory mice, guinea pigs, or rabbits.
Tick inoculation
Molted females of each tick species were allowed to rest in desiccators for a minimum of 30 days, and subsequently inoculated with about 1 μL virus suspension into the body cavity (hemolymph) through a coxal plate of the second pair of legs. Tick inoculation was performed using standard microcapillary tubes (Micropipette Puller P-30; Sutter Instrument Co., Novato, CA), connected to a microinjector (Digital Microinjector™ system MINJ-D-CE; Tri-Tech Research, Inc., Willoughby, OH).
Study design
The CYD-1–4 viruses, their DENV-1–4 and YF 17D parents, and the TBEV-positive control viruses, were each injected into fasting ticks. The inocula were titrated post-injection by plaque assay on Vero or on PS (TBEV) cells (Table 1).
Ticks were inoculated parenterally with 1 μL of each virus. The inocula were titrated for an accurate estimation of the injected dose. Each group was composed of 20 ticks inoculated with the same dilution of virus and incubated for a given period (2, 16, or 44 days) before feeding on four mice (Ixodes ricinus), or on one guinea pig (Rhipicephalus appendiculatus). Regarding TBEV, only one incubation time point (16 days) was tested, at the lowest inoculated doses (25 and 0.25 PFU).
After injection, the ticks were incubated at room temperature (∼23°C) for 2, 16, or 44 days in a desiccator (85% humidity). Control ticks inoculated with TBEV received three dilutions of virus and were tested after 16 days of incubation, except for the highest dose, which was tested after 2, 16, and 44 days. Following incubation, the ticks were fed on laboratory animals (I. ricinus: BALB/C mice; R. appendiculatus: guinea pigs). Small, round neoprene capsules with an opening on the top were glued onto the shaved backs of the animals using non-irritating latex glue. Five I. ricinus females/5 males, or 20 R. appendiculatus females/20 males of the same experimental group were placed together in each capsule, and the capsules were covered with nylon mesh. Each group was composed of 20 female ticks fed on four mice (I. ricinus), or 1 guinea pig (R. appendiculatus). After 3 days of feeding, the ticks were removed from the hosts, their salivary glands (SG) were dissected, and homogenates of both SG and body carcasses were prepared and titrated.
Transmission experiments
Forty-eight I. ricinus females (24 per virus) were inoculated with 3000 PFU of CYD-2 or 400 PFU of YF17D, and 12 tick females of each group were incubated for 16 or 44 days, respectively. As positive control, 20 I. ricinus females were inoculated with 2500 PFU TBEV and incubated for 16 days. Six mice were used for each group, except for TBEV (10 mice). In each experimental group, two inoculated female ticks co-fed for 3 days on individual BALB/C mice with 2 males and 15 uninfected nymphs. At the end of the feeding period, SG of inoculated females were dissected and stored at −80°C to check for infection, and homogenates of engorged nymphs were prepared and tested individually in plaque titration assays.
Preparation of tick homogenates
SG of 3-day-fed female ticks were dissected under a stereomicroscope under sterile conditions and rinsed twice in sterile 0.15 M NaCl. SG were placed individually in 50 μL sterile 0.15 M NaCl (TBEV-inoculated ticks), or in 100 μL RPMI medium/10% sorbitol/5% FBS/antibiotics (other inoculated ticks) and homogenized. The volume was filled to 500 μL with culture medium, the samples were centrifuged at 2000 rpm for 10 min at 4°C, and the supernatants were stored.
Adult tick carcasses (remaining after SG dissection) and tick nymphs were frozen at −80°C, then quickly thawed prior to use, and homogenized in 600 μL of RPMI medium/10% sorbitol/5% FBS/antibiotics (carcasses), or 500 μL of culture medium (nymphs), before centrifugation for 10 min at 2000 rpm and 4°C.
qRT-PCR analyses
Viral genomic RNA was quantified by qRT-PCR. Total RNA was extracted from 140 μL of tick homogenates (SG, body carcasses, or nymphs), using a Nucleospin 96 virus kit (Macherey Nagel, Düren, Germany), on a Tecan EVO150 robot (Tecan, Mannedorf, Switzerland), and eluted in 140 μL of nuclease-free water. PCR reactions were set-up with the one-step Quantitect Probe RT-PCR kit (Qiagen, Valencia, CA) in a final volume of 25 μL (0.5 μM of each primer, 0.2 μM of FAM-TAMRA probe, and 10 μL of RNA), and were run on an ABIPrism 7000TM (Applied Biosystems, Foster City, CA) thermocycler (50°C for 30 min; 95°C for 15 min; and 40 cycles of 95°C for 15 sec/60°C for 1 min). The primers and probes used in the YF NS5 qRT-PCR assays (quantification of both YF17D and CYD viral RNA) were previously published (Mantel et al. 2008). DENV-1–4 RNAs were quantified using serotype-specific primers and probes located in the NS5 gene: DENV-1: 5′-TCGCAACAGCCTTAACAG-3′/5′-ACTATCTCCCTCCCATCCTTC-3′, 5′-FAMTTCACACCACTTCCACTAMRA-3′; DENV-2: 5′-AATGACAGACACGACTCC-3′/5′-CCCAAAACCTACTATCTTCAA-3′, 5′-FAM TGGAAGTCGGCACGTGATAMRA-3′; DENV-3: 5′-AATGCGGAACCAGAAAC-3′/5′-ATTGCCATCTGTGTCACC-3′, 5′-FAMTCAGCCTCCTCCATGATAAATGTAMRA-3′; DENV-4: 5′-GTCAATCCACGCTCATCATC-3′/5′-TCCACACCACAAATCCTCTC-3′, 5′-FAMCAAAGTGTGGAACAGAGTGTAMRA-3′. RNA quantification standards were obtained as described previously (Mantel et al. 2008). Depending on the serotype, the low limit of quantification of the qRT-PCR assays was between 2.7 and 3.3 log10 GEQ/mL.
The TBEV genome was quantified using a previously published qRT-PCR assay (Schwaiger and Cassinotti 2003).
Statistical analysis
Analysis of variance (ANOVA) was used to assess the results, and data were processed with SAS PROC MIXED SAS v9.1 software, using a type I error risk equal to 10% for the interaction effects, and 5% for the main effects. Two fixed factors were included on the model, as well as the interaction (dose, day, and dose×day).
Results
Virus replication in tick salivary glands
I. ricinus or R. appendiculatus ticks were inoculated intracoelomically with about 3000 PFU of each CYD virus, and only about 300 PFU of DENV-1–4 and YF 17D, due to the lower titers of these lots. To allow comparison, CYD-2 was inoculated at two doses: 3000 and 300 PFU/tick. Actual injected doses were determined by titration of the inocula (Table 1). SG were dissected from the ticks after 2, 16, or 44 days of incubation and 3 days of feeding, and homogenized. Quantification of infectious virus or genomic RNA was performed on 80% or 10%, respectively, of each homogenate.
Ixodes ricinus
TBEV replicates in I. ricinus and was inoculated as a control, using three doses of virus (0.25, 25, and 2500 PFU). TBEV replication in I. ricinus SG was detected as soon as 2 days after inoculation in 7/8 ticks, and in 100% of inoculated ticks by day 16 at all doses, and at similar geometric mean titers (GMT) (3000–4000 PFU/tick). A 300-fold GMT increase was observed between day 2 and day 44 (a 10-fold increase was seen between days 16 and 44), in ticks inoculated with 2500 PFU (Table 2). In contrast, replication of parental DENV could not be detected in tick SG by either PFU or qRT-PCR assays (data not shown), and low levels of infectious YF 17D were detected in a maximum of 40% (day 16) of inoculated ticks (Table 3). No significant GMT increase was observed between days 2 and 16, and GMT tended to decline by day 44.
GMT was calculated for samples containing ≥1 PFU.
CYD-2-inoculated ticks in which all 20 ticks were analyzed.
Detection by PCR was performed on all tick salivary glands (20/group). Salivary glands from 10 ticks per group were tested in PFU assays.
NA, not applicable.
Infectious virus was detected at day 2 post-inoculation in SG of 60–100% of CYD-1–4-inoculated-ticks (Table 3). On day 16, 60% (CYD-1), 85–100% (CYD-2), 80% (CYD-3), and 20% (CYD-4) of all ticks were found to be positive. In at least 60% of ticks positive for CYD-1, CYD-3, and CYD-4, and about 25% of those positive for CYD-2 (regardless of exposure dose), fewer than 5 PFU were detected. Again in contrast to TBEV, no titer increase was observed between days 16 and 44 in ticks inoculated with either CYD-1–4 or YF 17D. By day 44, CYD-1, CYD-3, and CYD-4 infections were cleared, and YF 17D and CYD-2 infections were declining at both inoculated doses. Similar or slightly higher proportions of positive ticks were detected with qRT-PCR than with infectious titration (Table 3).
Compared with TBEV, genomic titers of CYD-1–4 and YF 17D were 300- to 10,000-fold lower (Fig. 1). Statistical analysis of individual values (n=20 per condition) revealed no significant differences between RNA titers measured at days 2, 16, and 44 after CYD-1–4 inoculation, other than significantly lower CYD-1 and CYD-3 genomic titers on day 44, compared with day 16 (p=0.014 and p=0.008, respectively).
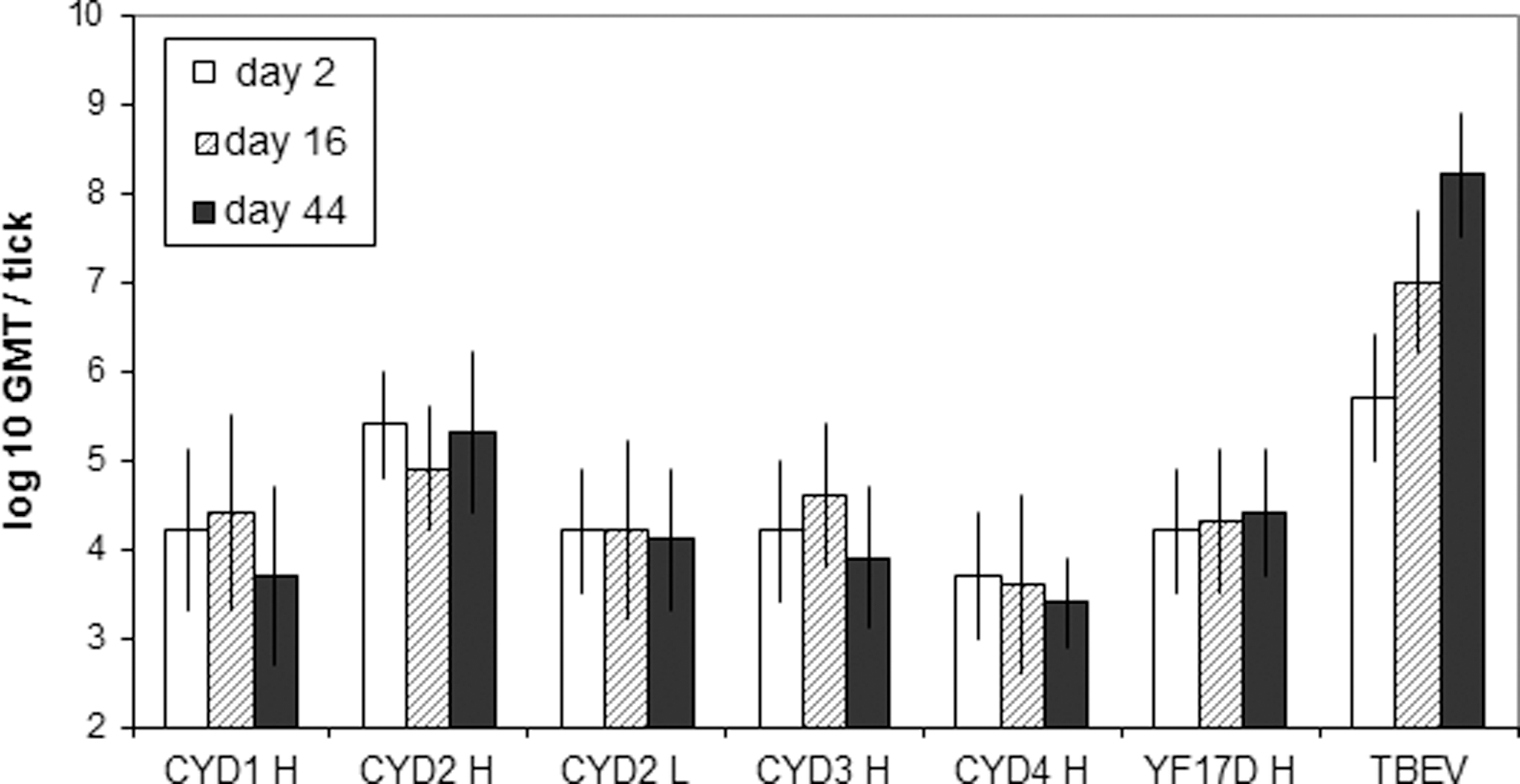
Quantification of total viral particles in salivary glands of Ixodes ricinus at different time points post-inoculation. Viral RNA was quantified by specific qRT-PCR in salivary glands of ticks inoculated with the higher (H), or the lower (L) dose of CYD-1–4 or YF17D viruses. For TBEV inoculation, ticks received 2500 PFU of virus. Each group represents the geometric mean titer (GMT) of 20 ticks.
Rhipicephalus appendiculatus
TBEV replication was not tested on R. appendiculatus, and DENV-1–4 did not replicate in this species. R. appendiculatus appeared less permissive than I. ricinus, whatever the virus tested (Table 4). By day 16, CYD-1 infection was already cleared, and infectious virus was detected in 60%, 10%, 80%, and 30% of ticks inoculated with CYD-2, CYD-3, CYD-4, and YF 17D, respectively. Overall, as was the case for I. ricinus, most detected samples (58%) contained <5 PFU. All viruses were cleared by day 44, except for 3 cases, for which SG samples contained 1 PFU. These trends were also observed by genomic RNA quantification (not shown).
GMT was calculated for samples containing ≥1 PFU.
NA, not applicable.
Virus detection in tick bodies
No replication of parental DENV was revealed in tick bodies. Infectious virus was detected in a maximum of 80% and 70% of I. ricinus tick carcasses (TC) inoculated with CYD-1 and CYD-2 viruses, respectively (Table 5). CYD-1 was cleared by day 44, while CYD-2 virus was still detected in 70% of cases. CYD-1 infection of R. appendiculatus ticks was detected in 40% of TC on day 16, and was cleared by day 44 (Table 5). RNA titers (Fig. 2) showed the same trends, and confirmed that infection of CYD-1 decreased in I. ricinus TC between day 16 and 44.

Quantification of viral genomic RNA in the bodies of Ixodes ricinus and Rhipicephalus appendiculatus (Ra) at different time points post-inoculation. Viral RNA was quantified by specific qRT-PCR in the carcasses of ticks inoculated with the higher dose of CYD-1 and CYD-2 viruses. Each group represents the geometric mean titer (GMT) of 20 ticks (GEQ, genome equivalents).
GMT was calculated for samples containing ≥1 PFU.
TC, tick carcasses without salivary glands; NA, not applicable.
Virus transmission through a non-infected host
Because CYD-2 virus persisted at low levels in I. ricinus ticks until day 44, transmission was tested in the co-feeding model. As controls, transmission of YF 17D and TBEV were also evaluated.
The CYD-2 and YF 17D viruses were detected in I. ricinus female SG inoculated with 3000 and 400 PFU, respectively, but negative results were obtained for all nymphs that co-fed with inoculated females (Table 6). In contrast, 64% (range 30–100%) of I. ricinus nymphs on average acquired TBEV infection in this model, and transmission occurred in each group that co-fed on individual mice. The average virus titer in positive nymphs was 98 PFU/nymph.
Positive ticks.
ND, not done.
Discussion
During the development of live genetically-modified vaccines, such as the CYD dengue vaccine candidate, one must ensure that they cannot propagate into the environment. The tissue/species tropism of the CYD chimeric viruses is driven by their dengue envelope, while their replication is driven by the YF17D polymerase (for a review see Guy et al. 2011). Previous studies have demonstrated that Aedes mosquitoes were unlikely to transmit CYD viruses (Johnson et al. 2002, 2004; Higgs et al. 2006). Here, the evaluation was taken a step further, and two hard tick species with different vector competence for flaviviruses and different geographical distributions were considered.
Ticks are not known to contribute to DENV circulation in the field (Labuda and Nuttall 2004), and no replication of DENV or of chimeric Langat/DEN4 or TBEV/DEN4 viruses in tick cell lines had been reported (Lawrie et al. 2004; Engel et al. 2011). Neither DEN4 nor chimeric Langat/DEN4 or TBEV/DEN4 viruses were able to infect or replicate in larvae of Ixodes scapularis, although these ticks are highly susceptible to infection with the tick-borne Langat virus (Engel et al. 2011). Accordingly, no viable DENV was detected in ticks at any point after inoculation in this study.
Although YFV has not been isolated from I. ricinus or R. appendiculatus in the field, data for other hard tick species indicate that YFV could persist and replicate in tick bodies, and even be transovarially and transstadially transmitted. However, there is no indication that ticks could be competent vectors of this virus in nature (Germain et al. 1979; Cornet et al. 1982; Davis 1993). In this report, we showed that YF 17D was not able to establish a robust infection in SG of I. ricinus or R. appendiculatus, in contrast to TBEV, in conditions that bypass the gut barrier and thus increase the susceptibility to infection.
Low levels of CYD-1–4 vaccine viruses were detected in I. ricinus SG on day 16 after direct inoculation, and with the exception of CYD-2 virus, which declined at a slower rate, they were cleared by day 44. Titers were highest in I. ricinus ticks inoculated with CYD-1 and CYD-2, but were in the same low range as those observed with YF17D virus, and at least 100-fold lower than those observed with TBEV. R. appendiculatus ticks were even less susceptible than I. ricinus to infection with CYD-1–4. No significant increase of infectious or genomic RNA titers over time was shown, indicating persistence rather than replication of these viruses. We cannot exclude hemolymph contamination of the tick SG extracts. We observed that while titers for CYD-1, CYD-3, and CYD-4 viruses were higher in carcasses than in SG of I. ricinus, the opposite was true for CYD-2. These results are in agreement with the actual presence of the latter virus in tick SG.
Non-viremic transmission of TBEV (i.e., transmission from infected to non-infected ticks co-feeding on the same hosts without detectable viremia) has been demonstrated and is considered to be an important mechanism for supporting virus circulation in nature (Labuda et al. 1993a, 1993b, 1996). As mice do not support replication of the CYD viruses, even after intracerebral inoculation, passive transmission between ticks was studied in such a model previously established for TBEV. Unlike TBEV, for which transmission occurred in 100% of the tested mice and in more than 60% of uninfected co-feeding I. ricinus nymphs, CYD-2 and YF17D were not detectably transmitted. Thus, while CYD-2 persisted to some extent in ticks after artificial inoculation, no transmission occurred, making such an event even less likely after vaccination.
To conclude, low-level, short-lived persistence of the four serotypes of CYD dengue vaccine viruses was observed after direct inoculation in a highly susceptible model of tick infection, which may reflect a self-limited low background replication, or simply represent persistence of the inoculum. Furthermore, no transmission was observed in this model. Together with the undetectable to low level of viremia observed after vaccination of human volunteers with tetravalent CYD vaccine (Morrison et al. 2010), these data demonstrate that the dissemination of these vaccine viruses by ticks after deployment of vaccine in the field is highly unlikely.
Footnotes
Acknowledgments
The technical assistance of Mrs. Zuzana Gulášová and Mrs. Marta Siebenstichová from the Institute of Virology SAS is gratefully acknowledged. The authors also wish to thank A. de Montfort and C. Hessler for performing the statistical analyses.
Author Disclosure Statement
This study was supported by Sanofi Pasteur, which is developing a dengue vaccine. N.M., S.R., J.L., B.G., and V.B. are employees of Sanofi Pasteur.