
Abstract
In this study, we performed a phylogenetic analysis of 35 species of the tribe Triatomini by means of available 16S ribosomal DNA and cytochrome b (Cyt b) gene sequence data, adding taxa of the spinolai complex, to clarify phylogenetic relationships of this complex and related triatomines. The phylogenetic analysis suggests a monophyletic clustering of the spinolai complex related to the South American species of triatomines.
Introduction
Until 1998, Mepraia spinolai was the only species of the genus, distributed in coastal and interior valleys from Chile between parallels 18° and 34° S (Lent and Wygodzinsky 1979). However, on the basis of karyotype, morphological characters, and experimental crosses, coastal desert populations between parallels 18°and 26° S were ranked as a new species named Mepraia gajardoi (Frías et al. 1998). The remaining populations, from 26° to 34° S, distributed in the interior mountains from the Atacama to the Metropolitan Regions, maintain the name M. spinolai (Frías et al. 1998). Recently, genetic studies using nuclear and mitochondrial markers on Mepraia sp. populations suggest the possible existence of introgression due to past hybridization events or the retention of ancestral polymorphisms, in the geographical intersection zone between these two taxa (Calleros et al. 2010). The goal of our study is to clarify the systematic uncertainties of Mepraia sp. and the Argentinan T. eratyrusiformis and T. breyeri. To this end, we constructed a phylogeny of Triatominae species belonging to the Triatomini tribe (Lent and Wygodzinsky 1979) by means of currently available mitochondrial gene sequence data of 16S ribosomal DNA and cytochrome b (Cyt b), adding the missing sequence data of the spinolai complex.
Materials and Methods
Insect collection
Collection of M. gajardoi specimens was carried in the locality of Caleta Vitor (Arica and Parinacota Region, 18° 45′45″ S, 70° 20′34″ W), while the M. spinolai were collected in the locality of Til Til (Metropolitan Region, 33° 06′19″ S, 70° 55′53″ W). In addition, we obtained Triatoma eratyrusiformis specimens from Salinas de Bustos (30° 15′50″ S, 67° 36′23″ W), and Triatoma breyeri from Patquia Viejo (30° 2′36″ S, 66° 52′46″ W). Both locations belong to Department of Independencia, Province of La Rioja, Argentina. Insects were collected manually; then they were dissected in the laboratory and their legs were kept in 70% ethanol at −20°C.
Mitochondrial DNA extraction, amplification, and sequence alignment
For DNA extraction, legs from each specimen were cut and macerated in an Eppendorf tube and incubated overnight with protease K, followed by DNA extraction with the EZNA Tissue DNA® kit according to manufacturer's instructions. A 663-bp fragment of the mitochondrial Cyt b gene was amplified via PCR using the primers and conditions as described (Monteiro et al. 2003). Verification of successful amplification was assessed by agarose gel electrophoresis. The amplified fragment was sequenced using the PCR primers in an internal service information
Phylogenetic analyses
Maximum likelihood (ML) and maximum parsimony (MP) algorithms were used to infer phylogenetic trees. ML analysis was inferred using the online platform PhyML 3.0 (Guindon and Gascuel 2003). The best-fitting model of nucleotide substitution was selected using the Akaike information criterion implemented in the program JmodelTest 0.1.1 (Posada 2008). These results gave a best fit for the GTR+G (G=0.2060, −lnL=8298.59467) nucleotide substitution model. Nodal supports were estimated by the bootstrap method (Felsenstein 1985) with 1000 replicates using PhyML 3.0 (Guindon and Gascuel 2003). MP analyses were performed using PAUP* 4.0b10 (Swofford 2003) with the heuristic search option, TBR algorithm, and 1000 replicates for the bootstrap method (Felsenstein 1985). Trees were visualized using the FigTree v1.1.2 program, available at
Results and Discussion
The phylogenetic analysis showed a clustering for the members of the spinolai complex, with support of 69% for this node (Fig. 1), which is marginally significant according to Hillis and Bull (1993). Within the complex, M. gajardoi and M. spinolai show a close relationship with significant nodal support values, close to T. breyeri and T. eratyrusiformis. Considering this phylogeny, we suggest that the species of the spinolai complex form a monophyletic group and that Mepraia corresponds to a clearly established genus and the phylogenetic relationship between T. eratyrusiformis and T. breyeri with M. spinolai and M. gajardoi reframes the debate to include these species in a single genus as proposed (Hypsa et al. 2002). According to our phylogeny, it would not be appropriate to synonymize Mepraia with Triatoma because a clear node supporting the genus Triatoma is lacking. In fact, species from the genera Panstrongylus and Dipetalogaster are also included within the genus Triatoma, suggesting a polyphyletic origin. However, it is necessary to include more gene sequence data from other species of the genus Triatoma to perform a more complete phylogenetic analysis.
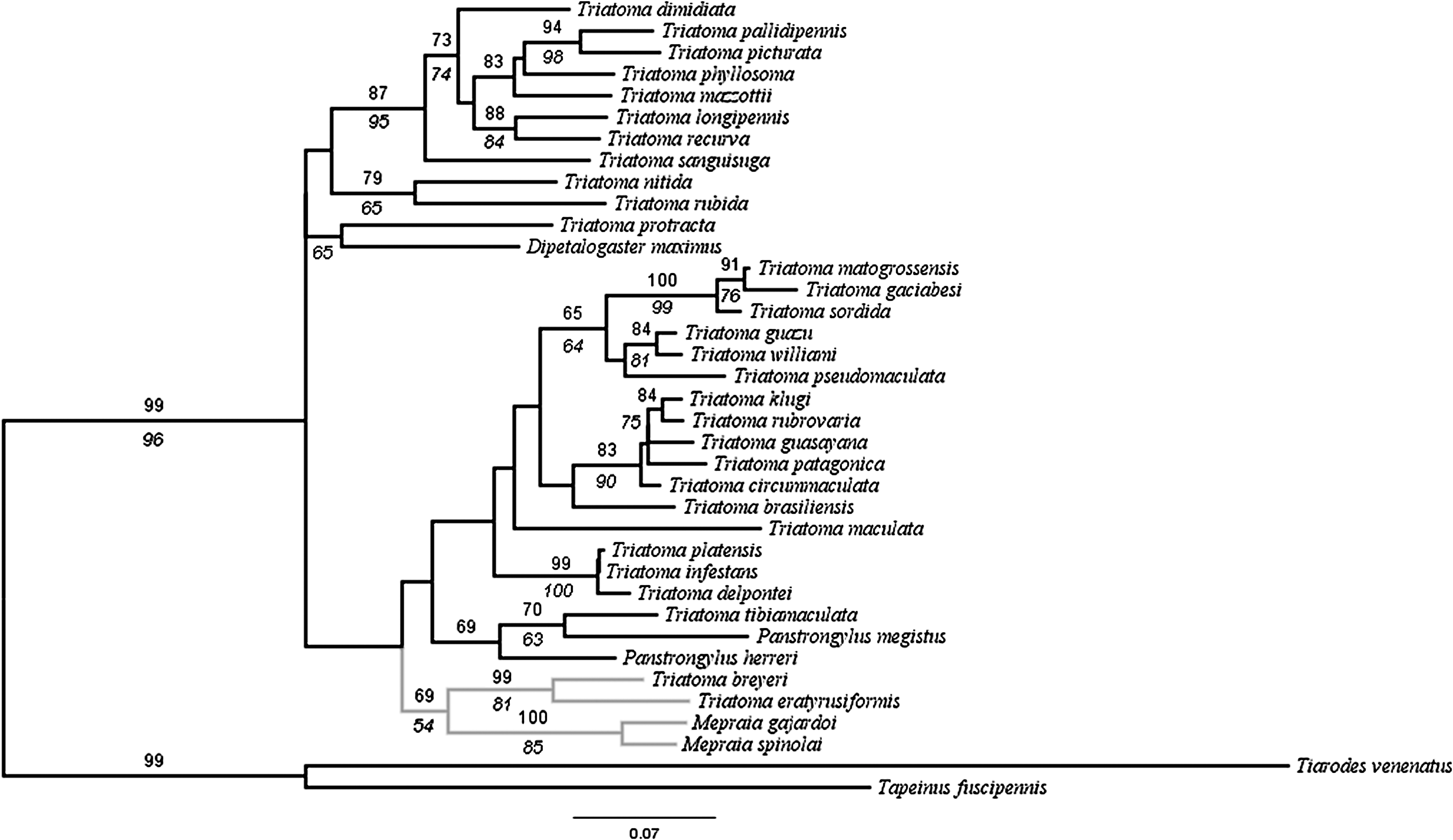
Maximum likelihood (ML) phylogenetic tree of Triatomini species based on 16S ribosomal DNA and Cyt b concatenated gene sequences, inferred with the model GTR+G (G=0.2060, −lnL=8298.59467). Numbers above the branch are the ML support values derived from bootstrap resampling with 1000 replicates and numbers under the branches are parsimony bootstrap values. Gray branches indicate the spinolai complex members.
The detected clustering of the spinolai complex members agrees with the classical studies proposed by morphological studies (Lent and Wygodzinsky 1979, Moreno et al. 2006). Moreno et al. (2006) compared the phenotype of the antennae among the species of the spinolai complex and found that Mepraia species and T. eratyrusiformis shared similarities in bristles and receptors that are not present in T. breyeri. In the same vein, it has been reported that T. eratyrusiformis shows multiple sex chromosomes like spinolai and gajardoi, but unlike most other South American Triatoma species (Panzera et al. 1998). It is necessary to complete the information provided by cytogenetic studies to have the whole scenario of the relationships of spinolai complex.
The monophyletic relationships between the taxa of the spinolai complex suggest that the Argentinian T. breyeri and T. eratyrusiformis could have had a common ancestor with the Mepraia genus. It has been proposed that the species of spinolai complex were separated with the uplifting of the Andes mountains during the Miocene in the Tertiary geological era about 20 million years ago (Moreno et al. 2006). It would be interesting to perform studies of the most recent common ancestor to estimate dates of divergence among the taxa to be compared with geological events. The spinolai complex appears related to South American species of triatomines. The relationship between M. spinolai and T. eratyrusiformis with South American species of the infestans complex were previously reported (Hypsa et al. 2002, de Paula et al. 2005). Interestingly, North American species seems to form a distictive group from South American species, even though not supported by high bootstrap values, which agrees with de Paula et al. (2005). This topology could be explained by the polyphyly of the Triatomini and the radiation pattern of these species (Schofield and Galvão 2009).
In conclusion, our phylogenetic evidence suggests that all the members of the spinolai complex constitute a monophyletic group corresponding to an established genus.
Footnotes
Acknowledgments
We thank Nicanor Villarroel (Universidad de Chile, Chile) and Natalia Folguera (CRILAR, Argentina) for laboratory assistance. Financial support was obtained from FONDECYT 1085154 (AS). Partial support was obtained from PBCT/PSD-66 and FONDECYT 11090086 (CBM). R Campos was supported by a CONICYT–PhD fellowship. S Catala is a member of CONICET, Argentina.
Author Disclosure Statement
No conflicts of interest are declared.