
Abstract
In malaria, mosquito saliva and salivary glands play central roles in the multi-faceted interactions that occur among the parasite, its vector, and its host. Analyzing the processes involved in the survival and maintenance of the Plasmodium parasite in mosquito organs, and in its transmission into vertebrate hosts, may lead to the identification of new molecular targets for parasite control. We used comparative two-dimensional gel polyacrylamide electrophoresis (2D-PAGE), surface-enhanced laser desorption/ionization time-of-flight mass spectrometry (SELDI-TOF-MS), and high-performance liquid chromatography (HPLC), followed by Edman sequencing, to study saliva and salivary gland samples from Anopheles gambiae mosquitoes infected or not with Plasmodium berghei. Quantitative 2D-PAGE profile analysis showed that the intensities of seven spots were affected by the presence of the parasite in the salivary glands. Most of the proteins identified possessed a signal peptide. SELDI-TOF-MS revealed 32 proteins/peptides whose peak intensities differed between the Plasmodium-infected and non-infected control groups. Quantitative comparison of HPLC profiles of low-molecular-weight components from salivary gland extracts revealed several peptides and proteins with levels that were modulated by parasite infection. The results of these complementary approaches suggest that the infection of female A. gambiae mosquitoes by P. berghei alters the production levels of several salivary gland proteins and peptides, some of which (e.g., protein cE5, B3VDI9_ANOGA, and AGAP008216-PA) are known or predicted to be secreted in saliva and involved in blood feeding.
Introduction
To interact with vertebrates, blood-feeding arthropods possess modified mouth parts and their salivary glands produce a highly adapted saliva containing components with pharmacological activities that counteract the hemostatic, inflammatory, and immune system responses of the vertebrate host (Ribeiro et al. 1984; Ribeiro 1995; Champagne 2005; Andrade et al. 2005; Titus et al. 2006).
Arthropod salivary glands play a crucial role in the development and transmission of vector pathogens. Indeed, in Plasmodium transmission, either by interacting with molecules within secretory cells (including saliva components), or because they undergo a time-programmed maturation, Plasmodium sporozoites in salivary glands are more infective to vertebrate hosts than earlier-stage sporozoites (Matuschewski 2006; Aly et al. 2009). Moreover, the pharmacological activity of arthropod saliva profoundly affects pathogen transmission (Ribeiro et al. 1987; Tabachnick 2000; Titus et al. 2006). Inoculation with Plasmodium via mosquito saliva increases the risk of infection, indicating that some parasites exploit the vector to enhance their development and multiplication in the vertebrate host (Rocha et al. 2004; Donovan et al. 2007). Using proteomic approaches to investigate the effects of Plasmodium sporozoites on salivary protein production is likely to provide insights into the migration and development of the parasite in a key organ for its maturation.
In a previous study, we used a number of complementary proteomic approaches to characterize the salivary gland proteome of A. gambiae (Choumet et al. 2007). In addition, we performed comparative analyses of Plasmodium-infected and non-infected salivary glands using two methods: a two-dimensional polyacrylamide gel electrophoresis (2D-PAGE)-based approach, in which the salivary gland extracts were subjected to a clean-up step, and a non-gel-based technique was used to quantify proteins (isobaric tags for relative and absolute quantitation: iTRAQ). Although we failed to identify any infection-specific markers with the 2D-PAGE-based method, the quantitative comparison performed by iTRAQ allowed us to identify five proteins that had levels that were modulated by Plasmodium infection (Choumet et al. 2007). Moreover, we did not address the putative modulation of salivary gland peptide production in the presence of the parasite. Indeed, several peptides were identified in the sialomes of Anopheles mosquitoes (Arca et al. 2005; Calvo et al. 2007), but the biological functions of most of these are unknown, especially with regard to their possible roles in Plasmodium infection. Salivary gland peptides may indeed play an important role in the cycle of the parasite and its transmission. Indeed, transcriptome analysis of Plasmodium-infected Anopheles stephensi showed a modulation of expression of several peptides (Dixit et al. 2009). Moreover, comparative analyses of transcriptomes from arbovirus-infected and non-infected arthropod salivary glands have shown induction of peptides with activity against some pathogens (Luplertlop et al. 2011; McNally et al. 2012).
In this study, to identify Plasmodium infection-specific markers, we performed comparative analyses of Anopheles salivary gland crude extracts using 2D-PAGE and surface-enhanced laser desorption/ionization time-of-flight mass spectrometry (SELDI-TOF-MS). We also analyzed the peptidic fraction of salivary gland extracts using ProteinChip array profiling. HPLC separation followed by sequencing of the peptides contained in infection-altered peaks allowed us to identify several Anopheles salivary gland peptide markers of P. berghei infection.
Materials and Methods
Mosquitoes and Plasmodium infection
A. gambiae mosquitoes were of a Yaoundé strain (Cameroon) and bred in the CEntre de Production et d'Infection des Anophèles (CEPIA) insectarium. Six-week-old Swiss mice (Janvier) were infected by intraperitoneal (IP) injection with a P. berghei strain NK65 expressing the green fluorescence protein (Natarajan et al. 2001). Parasitemia was monitored 4 days after Plasmodium injection in infected mice by examining Giemsa-stained thick blood smears. Mice exhibiting parasitemia at 7 days were considered to be infected. Three-day-old mosquitoes were fed on mice demonstrating 5–10% parasitized red cells in which gametocyte exflagellation was evident in a drop of blood mixed with PBS and viewed at 40× magnification. These same conditions were used for the seven infection experiments. Three-day-old control mosquitoes were fed on non-infected rabbit blood. The mosquitoes were maintained on a diet of 10% sugar.
Salivary gland collection
Salivary glands were collected from 21-day-old female mosquitoes, infected or not infected with P. berghei, 18 days after the blood meal. Infected salivary glands were identified using fluorescence microscopy. After dissection, the salivary glands were placed in a tube containing 100 μL of 150 mM sodium chloride (NaCl buffer), or a formate buffer consisting of 0.1 M formate (pH 3.5), and 150 mM NaCl with protease inhibitors. The samples were stored at −80°C until use.
Extract preparation
The salivary glands stored at −80°C were placed on ice for 1 h. They were sonicated using the following protocol: five runs of 3 min of 2-sec-long pulses at 100% amplitude. The extracts were then centrifuged at 130,000 g for 30 min at 4°C. The supernatants, designated salivary gland extracts (SGE), were stored at −80°C.
Determination of protein concentration and sample concentration
The protein concentration of SGE was determined spectrophotometrically using a ND-1000 Spectrophotometer (Nanodrop, Wilmington, DE). SGE samples were lyophilized, and either resuspended in 20 μL of sterile distilled water for immediate use, or stored dried at −80°C.
Two-dimensional gel electrophoresis
SGE samples from several infection experiments were pooled. Three samples of 15 μg lyophilized SGE were analyzed for each state: infected and non-infected, and corresponded to biological replicates. Samples containing 15 μg of SGE in a volume of 6.7 μL were placed on ice for 20 min, then vortexed and centrifuged for 5 sec. Then, Benzonase® (Sigma-Aldrich, St. Louis, MO) was added to a final concentration of 1.2%, and the samples were incubated for 1 min at 4°C. The samples were then mixed with 118.3 μL of rehydration solution containing 1% carrier ampholytes (pH 3–10; Invitrogen, Carlsbad, CA), 7 M urea (Bio-Rad, Hercules, CA), 2 M thiourea (Sigma-Aldrich), 4% CHAPS (Sigma-Aldrich), 100 mM DTT (Bio-Rad), and 0.002% bromophenol blue (Sigma-Aldrich). In-gel rehydration was performed using strip holders. The first-dimension separation protocol was conducted on 3–10 non-linear immobilins (7 cm) as follows: a passive rehydration for 16 h, an active rehydration for 15 min at 50 V, 30 min at 300 V, 30 min at 1000 V, and 2 h at 5000 V. The total run was approximately 6.5–7 kV-h with a maximum current of 50 μA/strip at 20°C (Ettan IPGphor II; GE Healthcare, Little Chalfont, U.K.). The strips were then equilibrated for 15 min in buffer containing 6 M urea (Bio-Rad), 50 mM Tris-HCl (pH 8.8; Sigma-Aldrich), 30% glycerol, 2% SDS, and 0.002% bromophenol blue, with the addition of either 1% DTT (Bio-Rad) for the first step, or 2.5% iodoacetamide (IAA; Sigma-Aldrich) for the second step. For the second dimension, the strip was loaded on a 1-mm-thick 12% SDS-PAGE gel without stacking gel, then embedded with 1% agarose. Gel electrophoresis was performed for 65 min at 120 mA/gel and 200 V. The slab gels were then stained with SYPRO Ruby (Invitrogen).
Analysis of gel patterns
SYPRO Ruby-stained gels were scanned with a Typhoon 9400 variable-mode imager (GE Healthcare), and the images were compared using ImageMaster 2D Platinum 7 software (GE Healthcare). All spots identified by the software were verified manually by eye. All spots reported were statistically significant according to the statistical tool built into the software (analysis of variance [ANOVA] test with p<0.05).
Protein identification by mass spectrometry
After SYPRO Ruby staining, all visible gel bands were excised using the robotic work station ProPic Investigator (Genomic Solutions, Ann Arbor, MI), and the plugs were collected into 96-well plates. The proteins were reduced, alkylated, and digested overnight with modified porcine trypsin (sequencing grade, ratio 1:100; Promega, Madison, WI) at 37°C using the ProGest Investigator (Genomic Solutions). The trypsin digests were desalted with C18 tips (μZipTip; Millipore, Billerica, MA). Peptides were directly eluted using the ProMS Investigator (Genomic Solutions) into the wells of a 96-well stainless steel MALDI target plate (Applied Biosystems/MDS SCIEX, Foster City, CA) containing 0.5 μL of α-cyano-4-hydroxy cinnaminic acid (CHCA; 2.5 mg/mL in 70% acetonitrile, 30% H2O, and 0.1% trifluoroacetic acid [TFA]).
Mass spectrometry analysis
Raw data for protein identification were obtained using a 4800 Proteomics Analyzer (Applied Biosystems/MDS SCIEX), and analyzed by GPS Explorer 2.0 software version 3.6 (Applied Biosystems/MDS SCIEX). For positive ion reflector mode spectra, 2000 laser shots were averaged. For MS calibration, autolysis peaks of trypsin ([M+H]+=842.5100 and 2211.1046) were used as internal calibrators. Monoisotopic peak masses were automatically determined within the mass range of 800–4000 Da with the signal-to-noise ratio minimum set to 30. Up to 25 of the most intense ion signals were selected as precursors for MS/MS acquisition, excluding common trypsin autolysis peaks and matrix ion signals. In the MS/MS positive ion mode, 4000 spectra were averaged; the collision energy was 2 kV, the collision gas was air, and the default calibration was set using Glu1-fibrinopeptide B ([M+H]+=570.6696) spotted onto 13 positions on the MALDI target. Combined peptide mass fingerprint (PMF) and MS/MS queries were performed using the MASCOT search engine 2.1 (Matrix Science Ltd., London, U.K.), embedded in the GPS Explorer software on the NCBInr database (downloaded on 01/19/2010; 10,348,164 sequences; 3,529,470,745 residues), with the following parameter settings: 50-ppm mass accuracy for MS; trypsin cleavage with one missed cleavage allowed; carbamidomethylation set as fixed modification; oxidation of methionines and formation of Pyro-Glu (N-term E and N-term Q) were allowed as variable modifications; and MS/MS fragment tolerance was set to 0.3 Da. Protein hits with a MASCOT protein score ≥83, and peptide hits with an Ionscore ≥53, each having a GPS Explorer protein confidence index ≥95%, were retained for further manual validation.
SELDI-TOF analyses
Supernatants of infected and non-infected salivary glands from various dissections were spotted on reverse-phase H4 (hydrophobic surface: C16), and weak cation exchanger (CM10: carboxylate) ProteinChip arrays (Bio-Rad). H4 chips retain peptides and proteins containing hydrophobic amino acids, whereas CM10 chips are designed to retain negatively-charged proteins. Prior to sample incubation, H4 chips were activated with 1 μL of 100% acetonitrile and air dried. SGE samples (1 μg) were spotted onto the H4 chips in duplicate, allowed to dry, and the spots were washed five times for 1 min each with 5 μL of MilliQ water. CM10 chips were equilibrated twice for 15 min with 150 μL of 100 mM sodium acetate (pH 3.8), using the Bioprocessor (Bio-Rad), and incubated with the same buffer containing 0.1% Triton X-100 for 20 min at room temperature. SGE samples (2 μg) were spotted onto the CM10 chips in duplicate and incubated for 30 min at room temperature with shaking (500 rpm). The arrays were washed under stirring for 5 min with binding buffer containing 0.1% Triton X-100, and then twice for 5 min with binding buffer without detergent. Then each spot was quickly washed twice with MilliQ water and allowed to air dry. Finally, 0.7 μL of a CHCA (BioRad) solution made by diluting a CHCA-saturated solution containing 50% acetonitrile and 0.5% TFA 1:5 in the same acetonitrile/TFA solution was loaded twice onto each spot of each chip (H4 and CM10) and the spots were air dried.
Molecules retained on the surfaces were visualized by reading the spots of each array in a ProteinChip System time-of-flight mass spectrometer (ProteinChip System 4000) using the ProteinChip MS reader software. Each array was read under suitable conditions for low mass (up to 20 kDa) to scan the spot, and shots were averaged at 810 nJ laser intensity with automatic data collection protocols in the Peaks (Ciphergen Biosystems Inc., Fremont, CA) software program. Low-mass spectra were calibrated on a mass calibration equation constructed with All-in-1 peptide standard mix of 7 (BioRad). The data obtained were analyzed using Ciphergen Express 3.0.6 software. Variant peaks were defined as those having p values<0.05.
Chromatographic comparison of salivary gland peptides
SGE from non-infected and infected mosquitoes were first separated by gel filtration on Superdex G75 columns (HR 3.3×30 mm) in formate buffer on an Ettan LC chromatography apparatus (GE Healthcare). Fractions (100 μL) containing peptides having molecular weights lower than 10 kDa were collected, then lyophilized and resuspended in 100 μL of 50 mM Tris HCl (pH 8.6) containing 0.01% Tween 20. Aliquots of 25 μL were then injected onto a DEAE column (150×1.1 mm) linked to a C18 reversed-phase HPLC column (150×1 mm), and eluted with a 2–70% acetonitrile gradient in 0.1% TFA. The profiles of the various fractions (both non-infected and infected) were then compared. Peaks having the same elution volume but differing by at least twofold in peak height between the two chromatograms, or being present in only one condition (infected or not), were then sequenced on a protein sequencer (ABI 494; Applied Biosystems). We conducted BLAST analyses to identify the peptides. The molecular weights of the eluted peptides were assessed by SELDI-TOF-MS on a Au ProteinChip array before sequencing.
Results
Comparative 2D-PAGE
In a previous 2D-PAGE analysis of desalted SGE (CleanUp kit; BioRad), we were unable to identify any salivary gland protein affected in the presence of P. berghei infection. We therefore analyzed non-desalted SGE (15 μg) on a non-linear pH 3–10 immobilin (7 cm) and designed an electrofocusing separation protocol. Three gels were performed for each condition (21-day-old infected and non-infected female mosquitoes). Differential analysis of the gels with Image Master 2D Platinum 7 revealed six spots that were upregulated and one spot that was downregulated (with a cut-off of 1.8-fold and ANOVA values of p<0.05) in the SGE from infected mosquitoes (Fig. 1, Supplementary Figure S1, and Supplementary Table S1; see online supplementary material at

2D-PAGE of infected salivary gland extracts. Extracts from salivary glands dissected 18 days after an infected or non-infected blood meal were separated by 2D-PAGE using pH 3–10 non-linear immobilins for the first dimension, and 12% acrylamide gel for the second dimension. Protein spots exhibiting differential intensities in infected salivary glands are indicated by number.
SELDI-TOF-MS comparative analysis of low-molecular-weight salivary gland components
SELDI-TOF-MS is a simple, rapid, high-throughput technique for profiling low-molecular-weight proteins and peptides, including those present in salivary glands. This technology is particularly well suited to identifying peptide markers of infection (De Bock et al. 2010). We examined two different chip surface chemistries: a weak cation exchanger (CM10), and a reversed-phase (H4) chip. Two dissection buffers were tested: 150 mM NaCl and 0.1 M formate in 150 mM NaCl (pH 3.5). The formate buffer was selected for its low pH, which decreases the strength of interactions between peptides and high-molecular-weight proteins. We detected multiple proteins having prominent peak intensities between 1000 and 18,000 m/z in infected and non-infected SGE. Figure 2 shows the profiles of infected and non-infected SGE dissected in formate buffer and separated on the ProteinChip surface (CM10). The selected proteins that varied in abundance with respect to infection status are listed in Table 2 according to their m/z ratio, the surface on which they were detected, and the dissection buffer used to prepare the extract. Altogether, the two different chip surfaces allowed us to identify 38 unique SGE polypeptides having molecular weights between 1067 and 18,917 Da, and had abundance levels that were affected by Plasmodium infection.

SELDI-TOF-MS spectra of non-infected and infected salivary gland extracts. The components of non-infected (
Variant peaks between non-infected and infected conditions were selected when the p value was<0.05. Peak molecular weights were obtained after calibration with molecular weight markers. Peaks characterized either on both surfaces or after using different dissection buffers are underlined.
Most of the peaks containing proteins that were less abundant in infected SGE than in non-infected SGE were identified using the CM10 chip, which proved to have the most efficient surface for identifying infection markers. We observed only a few overlaps for the resolved peaks (m/z) among the two array surfaces and the two dissection buffers, which indicated that the different surface chemistries resolved complementary protein spectra. The formate dissection buffer allowed a greater number of infection-altered peaks to be detected on the two chip surfaces. Thus we used extracts prepared from salivary glands dissected in this buffer to identify the proteins in these peaks in subsequent analyses.
Identification by HPLC of low-molecular-weight salivary gland components whose abundance is affected by P. berghei infection
Several A. gambiae salivary gland peptides whose abundance levels were modulated by P. berghei infection were identified using the SELDI procedure. However, since the amino acid sequences of these peptides could not be determined using this method, we analyzed them by HPLC and Edman sequencing. SGE proteins from non-infected and from P. berghei-infected female mosquitoes were first separated according to their molecular weight by gel filtration on Superdex G75, which generated different chromatograms for the two extracts (Fig. 3). Next, the non-infected fractions with molecular weight less than 10 kDa were compared to those of the SGE from infected mosquitoes, and individual peaks were compared between the extracts. The HPLC chromatograms obtained from the B5 fractions of infected and non-infected SGE separated on Superdex G75 show examples of three types of modulation (Supplementary Figure S2; see online supplementary material at
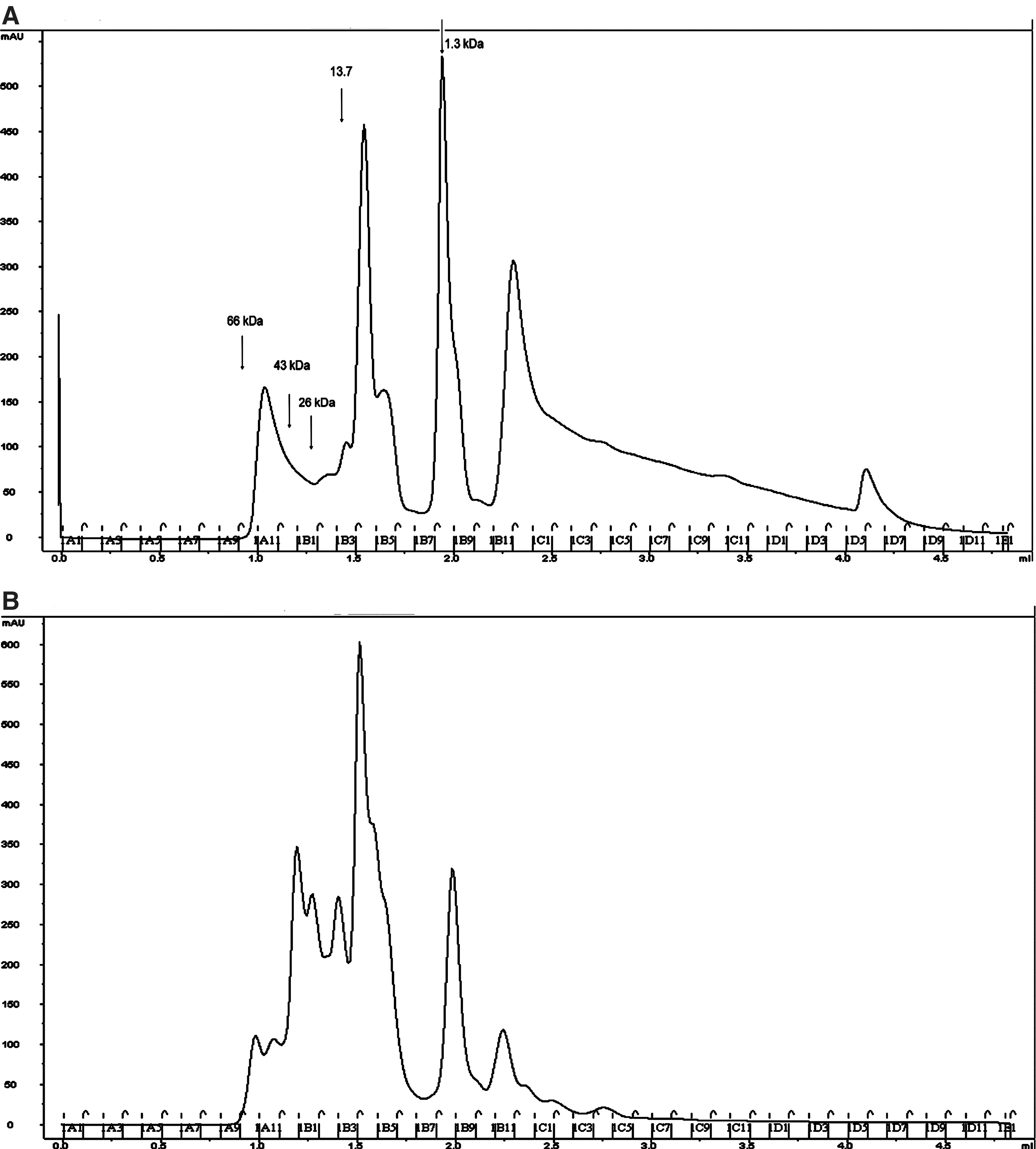
Separation of non-infected and infected salivary gland extract components by gel filtration. Salivary gland extracts from non-infected (
Modulation of the composition of salivary gland components was compared between infected and non-infected mosquitoes either from a pool of HPLC fractions smaller than 10 kDa, or by comparing individual fractions smaller than 10 kDa.
The fold modulation observed in infected/non-infected peptide is indicated for each experiment.
We found four markers representing three different proteins. One of these, the ce5 protein, was less abundant in infected salivary glands than in non-infected glands. Two peptides that were more abundant in infected salivary glands appeared to be fragments of the same protein: the 30-kDa salivary antigen of A. gambiae. This observation confirmed that this 30-kDa protein was the one in spot 4 of the 2D-PAGE differential comparisons that exhibited greater intensity in infected SGE than in non-infected SGE. The third peptide marker matched sequences found in the AGAP008216-PA protein, and in the gSG7 protein (with one amino acid difference).
Discussion
This work reveals elements of the various processes involved in the growth of Plasmodium parasites in mosquito salivary glands and in their subsequent transmission to a new host. Our approach identified new molecular targets that might be used for parasite control. A previous transcriptomic approach showed that 37 immune-related genes were upregulated in infected mosquitoes, as well as genes involved in protein transport, lipid metabolism, and energy metabolism. However, this approach indicated that the expression levels of genes implicated in blood feeding were only slightly modified (Rosinski-Chupin et al. 2007). The combination of comparative 2D-PAGE, SELDI-TOF-MS, and HPLC followed by Edman sequencing, allowed us to identify several putative protein and peptide markers. Several types of protein level modulation were detected in the presence of P. berghei infection. A. gambiae markers became more abundant, less abundant, disappeared, or appeared. The biological roles of some of these markers are unknown, including the salivary gland 1-like-3 (SG1-like-3) protein, a salivary gland protein of the G1 family of anopheline proteins (Arca et al. 2005). Transcripts of this protein family are found exclusively in or are enriched in the salivary glands of adult females, which suggest that the encoded proteins function in blood feeding. SG1-like-3 protein belongs to the same family as SG1b and TRIO proteins, which were also identified in spots of greater infection-specific intensity found in this study. Many of the other proteins we identified have known or putative functions; thus it may be possible to interpret the consequences of their modulation.
Markers involved in sporozoite entry or exit
Two proteins found to be in greater abundance in infected SGE were detected in spot 6 of the 2D-PAGE analysis: saglin and gSG1b. gSG1b was one putative Anopheles saliva protein recognized by sera from travelers in tropical Africa (Orlandi-Pradines et al. 2007). This result suggests that gSG1b is indeed a saliva protein; however, its role has not yet been determined. Saglin has been shown to be involved in the invasion of salivary glands by P. berghei sporozoites via its interaction with the TRAP protein (Okulate et al. 2007; Ghosh et al. 2009). Consequently, increased production of saglin might facilitate sporozoite entry into and exit from salivary glands.
Markers involved in blood feeding and parasite transmission
Some of the markers we identified, such as cE5, B3VDI9_ANOGA, and gSG6, are clearly involved in blood feeding, and their modulation may influence this behavior. The cE5 and gSG6 proteins were less abundant in infected mosquitoes, whereas the level of B3VDI9_ANOGA was higher in infected mosquitoes. B3VDI9_ANOGA is a member of the 30-kDa saliva antigen family. Transcripts encoding members of this acidic protein family were detected in a previously reported study of culicine and anopheline mosquito transcriptomes (Calvo et al. 2007). Anti-platelet activity has been attributed to some of the members of the SG1 family that are similar to B3VDI9_ANOGA. Protein cE5 is an analogue of anophelin, a peptide inhibitor of thrombin identified in A. stephensi (Francischetti et al. 1999; Valenzuela et al. 1999; Yoshida et al. 2008). Silencing of this gene using RNAi led to lower blood-feeding capacities and increased probing times (Das et al. 2010), which resulted in increased Plasmodium transmission.
The gSG6 protein levels were lowered by infection, which corroborates our earlier findings (Choumet et al. 2007). Injection of gSG6 dsRNA into adult A. gambiae females decreased gSG6 protein levels, increased probing time, and reduced blood-feeding ability (Lombardo et al. 2009), all of which contributed to higher levels of parasite transmission.
Another marker of infection, AGAP008216-PA, is homologous to the gSG7 protein. The sequence of the A. gambiae gSG7 protein is 63.4% identical to anophensin, which is a kallikrein-kinin system inhibitor from the salivary glands of another malaria vector mosquito, A. stephensi (Isawa et al. 2007). Anophensin strongly inhibits the release of bradykinin through inhibition of kallikrein-kinin system activation (Isawa et al. 2007). The anti-inflammatory activity of this protein might be useful to the parasite because Plasmodium-infected mosquitoes probe for longer times than non-infected mosquitoes, and are therefore more likely to attract the attention of the host (Wekesa et al. 1997). This may enhance the likelihood of parasite transmission.
Conclusions
By using complementary proteomics approaches, we confirmed that P. berghei sporozoites were able modify the compositions of salivary glands and saliva by affecting either gene expression, or the rate at which various proteins are metabolized. Most of these effects would appear to favor parasite penetration, transport, or transmission. The identification of new infection markers will improve our understanding of the complex and crucial relationship between the parasite and the salivary glands of vector mosquitoes. This in turn could lead to new strategies for preventing malaria transmission by targeting its vector.
Footnotes
Acknowledgments
The work was supported by grants from the Institut Pasteur (Grand Programme Horizontal Anophèle), CNRS-PAL+, and the Research Program on Parasitic Diseases Sanofi-Aventis/Ministry for Research. We thank the CEPIA of Institut Pasteur (Dr. Catherine Bourgouin) for mosquito breeding and Plasmodium berghei infections. We thank Dr. Charles Roth and Dr. Paul Brey for their support and advice during this work, and for critical reading of the manuscript.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.