
Abstract
Rickettsiae are obligate intracellular bacteria that cause zoonotic and human diseases. Arthropod vectors, such as fleas, mites, ticks, and lice, transmit rickettsiae to vertebrates during blood meals. In humans, the disease can be life threatening. This study was conducted amidst rising reports of rickettsioses among travelers to Kenya. Ticks and whole blood were collected from domestic animals presented for slaughter at major slaughterhouses in Nairobi and Mombasa that receive animals from nearly all counties in the country. Blood samples and ticks were collected from 1019 cattle, 379 goats, and 299 sheep and were screened for rickettsiae by a quantitative PCR (qPCR) assay (Rick17b) using primers and probe that target the genus-specific 17-kD gene (htrA). The ticks were identified using standard taxonomic keys. All Rick17b-positive tick DNA samples were amplified and sequenced with primers sets that target rickettsial outer membrane protein genes (ompA and ompB) and the citrate-synthase encoding gene (gltA). Using the Rick17b qPCR, rickettsial infections in domestic animals were found in 25/32 counties sampled (78.1% prevalence). Infection rates were comparable in cattle (16.3%) and sheep (15.1%) but were lower in goats (7.1%). Of the 596 ticks collected, 139 had rickettsiae (23.3%), and the detection rates were highest in Amblyomma (62.3%; n=104), then Rhipicephalus (45.5%; n=120), Hyalomma (35.9%; n=28), and Boophilus (34.9%; n=30). Following sequencing, 104 out of the 139 Rick17b-positive tick DNA had good reverse and forward sequences for the 3 target genes. On querying GenBank with the generated consensus sequences, homologies of 92–100% for the following spotted fever group (SFG) rickettsiae were identified: Rickettsia africae (93.%, n=97), Rickettsia aeschlimannii (1.9%, n=2), Rickettsia mongolotimonae (0.96%, n=1), Rickettsia conorii subsp. israelensis (0.96%, n=1), Candidatus Rickettsia kulagini (0.96% n=1), and Rickettsia spp. (1.9% n=2). In conclusion, molecular methods were used in this study to detect and identify rickettsial infections in domestic animals and ticks throughout Kenya.
Introduction
Despite the availability of doxycycline, a cheap and effective treatment for rickettsioses, many cases go untreated as “fevers of unknown origin” largely due the lack of obvious clinical symptoms in the local population, and the overwhelming cases of malaria infections that may further mask rickettsial infections (Rutherford et al. 2004). On introduction to a human host, the rickettsial pathogens multiply in the vertebrate endothelial cells and, due to ensuing vasculitis, may produce an eschar at the site of the tick bite that may be useful in diagnosis. Other symptoms include fever, headache, myalgias, and rash. Rarely, papulovesicular lesions may resemble rickettsialpox. Regional adenopathy, disseminated lesions, kidney disease, splenic rupture, and focal hepatic necrosis may occur (Parola et al. 2005).
The genus Rickettsia is divided into 3 main biogroups (La Scola and Raoult 1997; Graves et al. 1999): (1) the SFG, which is largely transmitted by ixodid ticks with exception of R. felis and R. akari, whose vectors are fleas and mites, respectively; (2) the TG, which is transmitted by fleas and lice; and (3) the scrub typhus group (STG) with chiggers as the main vectors.
Two SFG rickettsiae have been identified in Africa, Rickettsia conorii and R. africae. R. conorii, is the etiologic agent of Kenyan tick typhus, also known as Mediterranean spotted fever (MSF). It is transmitted predominately through the bite of infected Rhipicephalus sanguineus tick (Raoult and Roux 1997). R. africae, the causative agent of Africa tick-bite fever is said to be largely transmitted by Amblyomma ticks. It has been reported in South and Central Africa, including Kenya (Kelly et al. 1996; Macaluso et al. 2003).
Screening for rickettsiae by molecular methods is often based on the 17-kD gene (htrA) that is specific for the Rickettsia genus (Anderson et al. 1987). For species identification, a combination of PCR and restriction fragment-length polymorphism targeting the rickettsial outer membrane proteins A gene (ompA) can be conducted on 17-kD positive samples (Eremeeva et al. 1994; Moncayo et al. 2010). Other gene targets for discrimination between different species within the genus target genes, such as rickettsial outer membrane protein B (ompB) and the citrate-synthase encoding gene (gltA). This study used molecular surveillance to describe rickettsial geographical distribution in Kenya, infection rates in domestic animals and ticks, and the Rickettsia species within the tick populations.
Material and Methods
Ethical consideration
This study was carried out using blood samples from domestic animals presented for slaughter. The study was conducted under protocol #1248 that was reviewed and approved by the Animal Use Committee of the Kenya Medical Research Institute.
Sample collection
Ticks and whole blood were collected from domestic animals presented for slaughter to major slaughterhouses in Nairobi (Athi River Kenya Meat Commission [KMC] slaughter house) and Mombasa (KMC Kibarani, Uwanja wa Ndege, Mariakani and Kasemeni) between November, 2007, and September, 2008. These slaughterhouses receive animals from nearly all counties in the country. Animal movement permits issued by the local veterinarian from the animal's area of origin were used to determine the animals' county of origin. Only a sample set of animals that were presented was sampled, and because there was a huge difference in the number of animals brought from different counties, sampling was not random. Efforts were made to include animals from as many counties as possible.
Blood was drawn antemortem, just before the animals were shot with a stun gun. EDTA blood was collected from each animal by jugular venipuncture. The blood was first centrifuged at 1500×g for 10 min in a benchtop centrifuge, and the buffy coat was carefully removed and stored in duplicate in 1.5-mL cryovials. The samples were stored at −70°C until required for analysis. At least 10 adult ticks were also collected from each animal found to have ticks. The collected ticks were placed in 1.5-mL vials containing 70% ethanol and transported to the laboratory.
Tick taxonomy
Ticks were identified taxonomically using the standard taxonomic key. Ticks of the same species and from the same domestic animal were pooled together and placed in a 1.5-mL vial containing 70% ethanol and stored at −70°C. Each vial containing classified pooled ticks was labeled with information of the tick species and identification of the tick's animal host.
DNA extraction
A total of 4 mL of EDTA blood was centrifuged, 200 μL of the buffy coat collected, DNA extracted using the QIAamp DNA Mini Kit (Qiagen Inc., Valencia, CA), eluted in 200 μL, and stored at −20°C until required. Ticks were removed from the 70% ethanol, allowed to rehydrate in 3 mL of phosphate-buffered saline (PBS) (pH 7.4) for 5 min. Pools of ticks from the same animal were homogenized in a mortar and resuspended in 1 mL of PBS. DNA was extracted from 200 μL of the homogenate using a Qiagen QiaAmp DNA Mini Kit according to manufacturer's instructions (Qiagen Inc., Valencia, CA), eluted in 200 μL, and stored at −20°C until required.
Screening and identification of Rickettsia
Host and tick DNA were screened for evidence of rickettsia using a quantitative real-time PCR (qPCR) assay (Rick17b) that amplifies and detects a fragment of the 17-kD gene essentially as described before (Jiang et al. 2012). For species identification of rickettsiae DNA in the tick DNA preparation, a single-step conventional PCR targeting the ompA, ompB, and gltA genes was carried out on Rick17b-positive samples using previously described primer sets (Regnery et al. 1991, Jiang et al. 2005). Four microliters of tick DNA was used as template in a 21-μL reaction mix containing GeneAmp PCR buffer, 200 μM of deoxyribonucleotide triphosphates (dNTPs) mix with deoxythymidine triphosphate (dTTP), 0.5 μM, 0.3 μM, and 1.0 μM of ompA, ompB, and gltA primer pairs, respectively, and 0.3 U/μL of AmpliTaq Gold (all from, Applied Biosystems, NJ). The reactions were run in a Biometra T3000 Thermocycler (Horsham PA) under the following conditions: For ompA, and gltA, 95°C for 3 min, followed by 35 cycles of 95°C for 30 s, 50°C for 30 s, 72°C for 30 s, and final extension for 7 min at 72°C. For ompB, the amplification conditions were 95°C for 5 min, followed by 35 cycles of 95°C for 15 s, 54°C for 15 s, 72°C for 30 s, and final extension for 7 min at 72°C. PCR products were separated on a 1.5% agarose gel, which was run in Tris-acetate-EDTA (TAE) electrophoresis buffer and detected by staining with 0.5 μg/mL ethidium bromide. The samples that did not amplify with the single-step conventional PCR were amplified by nested PCR, as previously described (Jiang et al. 2005).
DNA sequencing and analysis
The ompA, ompB, and gltA PCR amplicons were purified using a QIAquick PCR purification kit as recommend by the kit manufacturer (Qiagen Inc.). Both strands of purified PCR products were sequenced using the Big Dye Terminator Kit (Applied Biosystems, Foster City, CA) and the sequence analyzed in a 3130 Genetic Analyzer (Applied Biosystems). Consensus nucleotide sequences minus the primers were used to query the GenBank database, and the highest similarity was identified by Basic Local Alignment Search Tool (BLASTN), available from the National Center for Biotechnology Information (Bethesda, MD).
Results
Rickettsia infection in domestic animals
A total of 1697 domestic animals comprising 1019 cattle, 379 goats, and 299 sheep were sampled between November, 2007, and September, 2008, from 32/47 counties in Kenya. The origin of the domestic animals was verified from animals' movement permit issued by the local veterinarian at the county of origin. There were no domestic animals presented for slaughter from 15 counties. Rickettsia infection rates in domestic animals as determined by the Rick17b qPCR assay were found in 25 out of the 32 counties (78.1%) sampled (Fig. 1). Rickettsial infection rates were comparable in cattle (166/1019, 16.3%) and sheep (45/299, 15.1%) and were lowest in goats (27/379, 7.1%). The difference was statistically significant (p<0.001).
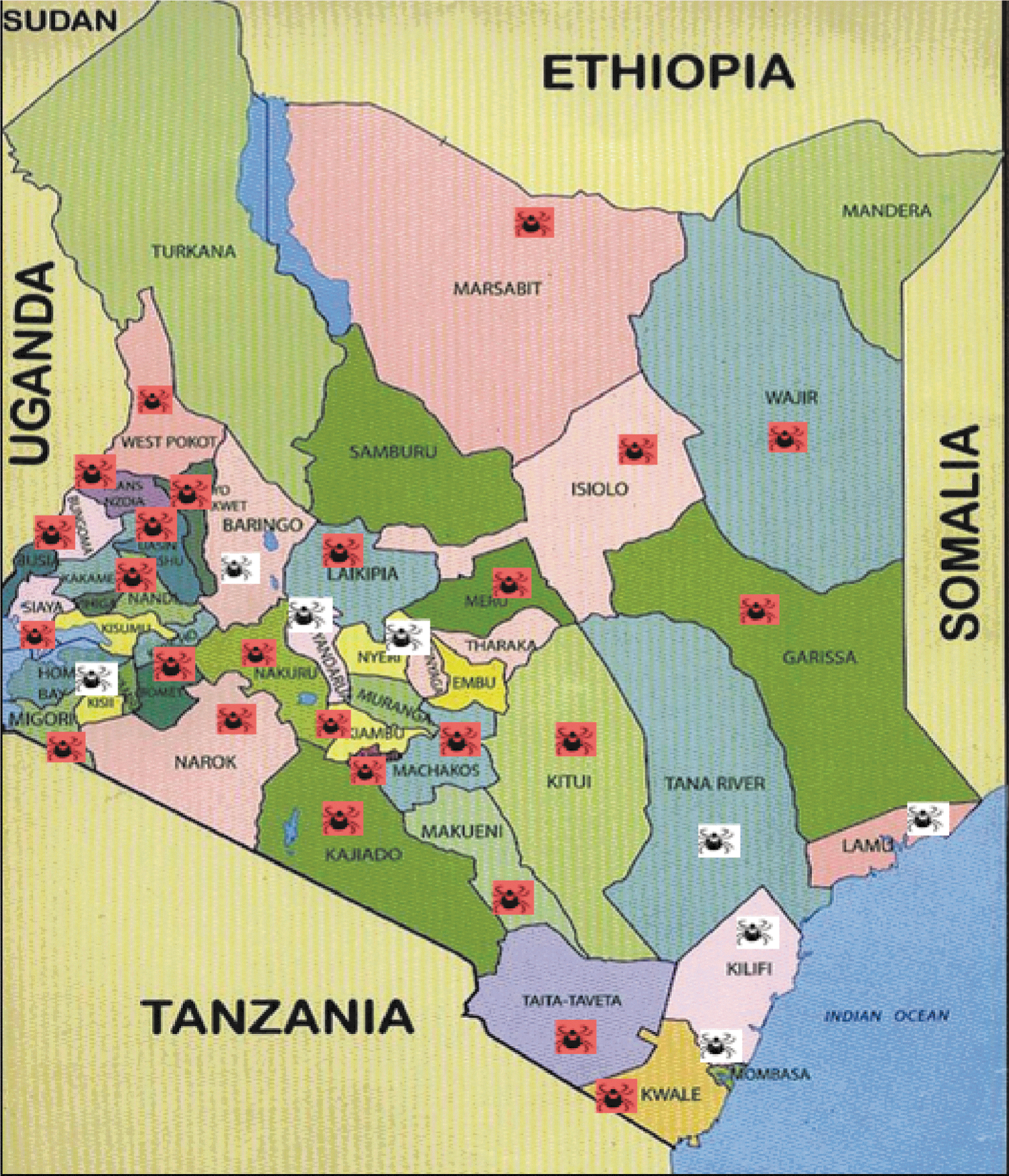
Map of Kenya showing counties where domestic animals were sourced and percent Rickettsia infection rates.
Tick species and infection rates
A total of 596 adult ticks were collected from domestic animals. Tick infestations were highest in cattle (72.9%) and sheep (22.5%) and lowest in goats (4.6%), and the differences were statistically significant (p<0.001). The ticks were taxonomically identified to belong to 4 tick genera: Amblyomma, Rhipicephalus, Hyalomma, and Boophilus. Ticks of the genera Rhipicephalus (264/596) and Amblyomma (167/596) were the most abundant in domestic animals at 44.3% and 28.0%, respectively, compared to Boophilus (14.2%) and Hyalomma (13.3%) (Table 1). Of the 596 adult ticks collected, 139 were found infected with Rickettsiae. Rickettsia infections were highest in Amblyomma (62.3%), followed by Rhipicephalus (45.5%), with Hyalomma and Boophilus at 35%. At the species level (Fig. 2), infection rates were similar among the Amblyomma species, except for the unspeciated Amblyomma (38.5%) compared to ≥60% for A. gemma, A. hebraeum, and A. variegatum. Among the genus Hyalomma, the unspeciated Hyalomma spp. had the lowest infection rate (6.7%) compared to 54.6% for H. truncatum. In Rhipicephalus, Rh. appendicutus had the highest infection rate (55%) compared to (38.7%) for the Rh. pulchellus.

Tick species, animal host, and percent of Rickettsia infection.
Identification of Rickettsia spp.
Of the 139 ticks that were found infected following the initial screening with the Rick17b assay, 104 (74.8%) amplified with all the 3 gene targets (ompA, ompB, and gltA; Fig. 3) and were sequenced; 35 (33.6%) others amplified with 1 or 2 of the 3 genes and were not sequenced. The reverse and forward consensus sequences for each target gene were used to query GenBank. GenBank accession numbers and percent identity are shown in Table 2. Six Rickettsia species belonging to SFG were identified (Table 2 and Supplemental Table S1 online at
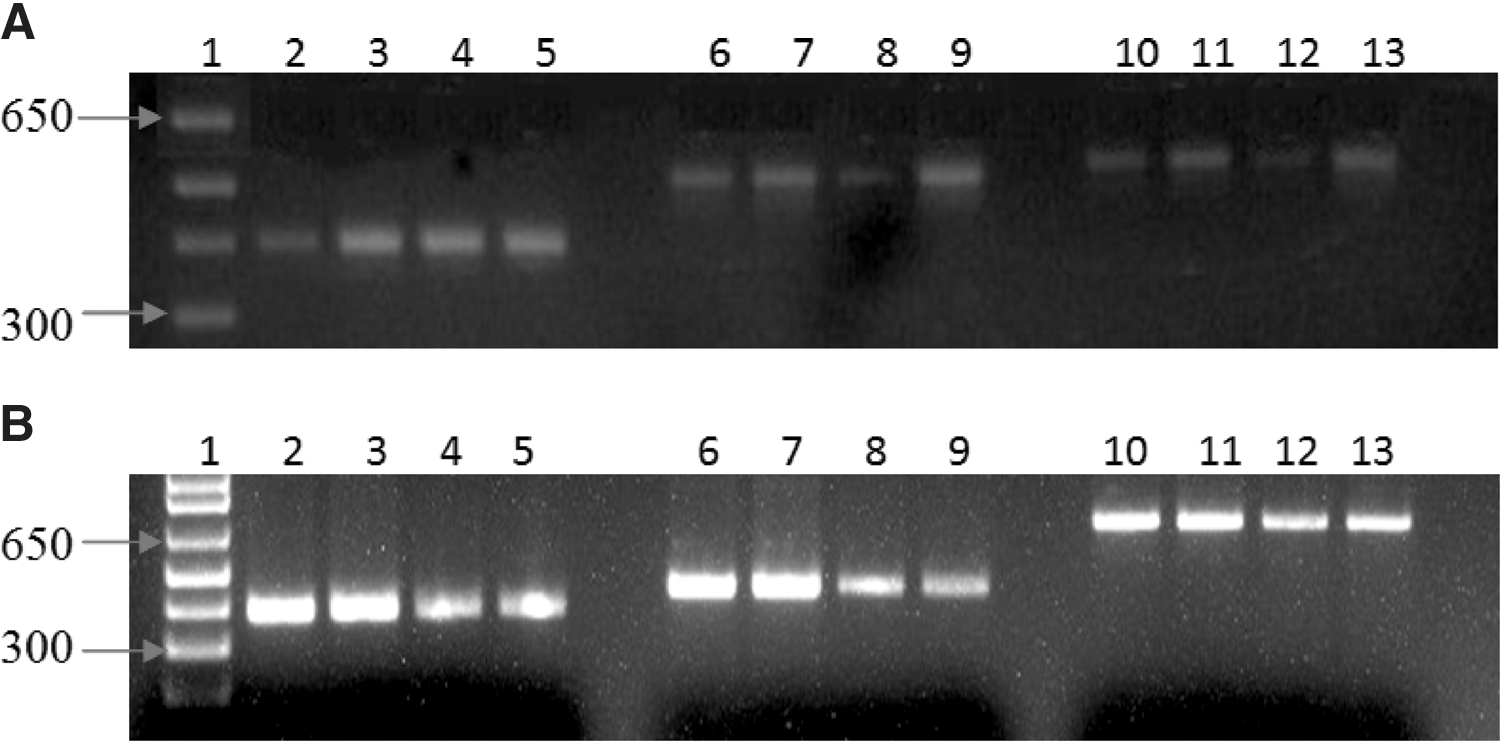
Typical amplification products for tick DNA samples using rickettsial-specific ompA, ompB, and gltA primers by single-step conventional PCR (
Failed, sequence failure; NA, not available in GenBank; ND, not determined because % identity was low for definite species identification.
The CLUSTAL multiple sequence alignments of the species identified are shown in Table 2. R. conorii subsp. israelensis did not amplify with ompA primers. The percent identity of the Rickettsia species to reference sequence ranged between 93% and 100% (Table 2).
Discussion
Rickettsioses are increasingly being reported in sub-Saharan Africa. In Kenya, a number of studies have over time identified various Rickettsia spp., including Rickettsia typhi (murine typhus) (Heisch 1969), R. africae (Macaluso et al. 2003), and more recently R. felis (Richards et al. 2010), illustrating the growing importance of this hitherto underrated zoonosis. The current study represents an effort to describe rickettsial epidemiology in Kenya.
In this study we used a nonprobability sampling technique to obtain pilot data on rickettsial epidemiology using domestic animals presented at slaughterhouses in Nairobi and Mombasa as source material. As shown in Fig. 1, 25/32 counties sampled had rickettsiae, illustrating how widespread this zoonosis is in Kenya.
Tick infestation was highest in cattle (72.9%) and sheep (22.5%) and lowest in goats (4.6%). The rickettsial infection rates were lowest in goats (7.1%, n=27) compared to cattle (16.3%, n=166) and sheep (15.1%, n=45). Because there was no difference in infection rates between cattle and sheep, despite a more than 3-fold difference in tick infestation, the low infection rates in goats were attributed to forage preferences between the species. The ticks of the genus Ambylomma and Rhipicephalus were the dominant carriers of rickettsiae, with infection rates of 62.3% and 45.5% respectively, compared to 35.9% in Hyalomma and 34.9% in Boophilus (Table 1, Fig. 2). This is consistent with earlier findings that implicated Amblyomma and Rhipicephalus as the main vectors of rickettsiae in Africa (Parola et al. 2001).
We used three rickettsial gene targets, ompA, ompB, and gltA, to query GenBank for each of the tick DNA that was amplifiable (Fig. 3 and Table 2). The use of these genes together gives better reliability in phylogeny assignments of the genus Rickettsia (Roux and Raoult 2000). All Rickettsia species identified were members of the SFG that cause “tick typhus.” R. africae, the causative agent of Africa tick-bite fever, was present in 94.2% of the ticks isolated, attesting to the success of this bacteria in African ticks. In humans, the bacterium causes fever and eschar formation at the site of the tick bite. R. aeschlimannii was present in 1.9% of the ticks. In addition to fever and eschar formation, R. aeschlimannii is associated with maculopapular rash. This the first report of R. mongolotimonae in Kenya. R. mongolotimonae is associated with an even more serious disease that is characterized by fever, multiple eschars, regional adenopathy, lymphangitis, and maculopapular rash. This is the first report of R. conorii subsp. israelensis in Kenya. In humans, the bacterium is associated with Israel spotted fever.
There is a strong geographical correlation between the prevalence of the Ambylomma and Rhipicephalus tick species and incidence of R. africae (Fournier et al. 1998b). In this study, A. gemma and A. hebraeum were the main vectors of R. africae. This is consistent with earlier work by Fournier et al. (1998a) and Kelly et al. (1991). Nevertheless, R. africae has been identified and isolated before from Rhipicephalus both in Kenya and the Central African Republic (Tissot-Dupont et al. 1994, Macaluso et al. 2003). In this study (Table 2), Rh. pulchellus and Rh. appendiculatus were also found infected with R. africae. In Africa, R. aeschlimanii has been isolated in the tick genus Hyalomma and Rh. appendiculatis (Beati et al. 1995, 1997, Pretorius and Birtles 2002). In our study, R. aeschlimannii was isolated from H. truncatum and B. annulatus. B. annulatus was found to be infected with Candidatus Rickettsia Kulagini. R. mongolotimonae was first reported in South Africa and isolated from the H. truncatum (Parola et al. 2001). In our study, R. mongolotimonae was isolated from A. gemma ticks.
From a recent study, SFG and TG rickettsiae form a significant part of undifferentiated febrile illness in hospitalized patients in Tanzania, but unfortunately these infections go undiagnosed and untreated (Prabhu et al. 2011). Rickettsioses in Africa are transmitted by a variety of vectors, including ticks, mites, lice, and fleas that have a number of hosts, including livestock, dogs, cats, and rodents. Vector control is unlikely to impact rickettsial infections in Africa due to a number of reasons, among them, the diverse ecology inhabited by the vectors, lack of appreciation of the morbidity attributable to rickettsioses, as well as social economic factors. Mitigation of rickettsioses in East Africa will likely come from increased awareness about this disease, early diagnosis, and prompt treatment with effective antibiotics, such as doxycycline.
Limitation of this study includes the inability to sample animals randomly, and therefore some counties were underrepresented. In Kenya, the beef industry is controlled by pastoralists and commercial ranchers, and both are located in the semiarid parts of the country. Therefore, more animals came from counties in the semiarid areas. Deliberate efforts had to be made to include whatever few livestock that came from counties outside the semiarid parts of the country. In general, commercial ranchers have better tick control programs than the pastoralist and, therefore, livestock from commercial ranches had fewer or no ticks compared to livestock owned by the pastoralists. Finally, certain ticks such as Rhipicephalus that are not considered competent vectors of rickettsiae were shown to harbor these pathogens. Without demonstrating infection in the ticks' lymph, it is difficult to claim that biological development was occurring in some of these ticks.
In conclusion, the molecular methods used in this study identified a variety of rickettsiae in domestic animals and various species of ticks. The obvious next move will be to initiate serological and molecular studies to determine rickettsial prevalence and burden in humans.
Footnotes
Acknowledgments
We are grateful to the Director of Veterinary Services for authorizing and facilitating conduct of this study. We thank project staff, including animal attendants and laboratory technicians. This work is published with the permission of the Director, Kenya Medical Research Institute. James Wainaina was a M.Sc. student at Jomo Kenyatta University of Agriculture and Technology. Financial support was from a Grant from the Global Emerging Infection System (GEIS).
The U.S. Government has the right to retain a nonexclusive, royalty-free license in and to any copyright covering this paper. The opinions or assertions contained herein are the private views of the authors, and are not to be construed as official, or as reflecting true views of the Department of the Army, the Department of the Navy or the Department of Defense.
Author Disclosure Statement
No competing financial interest exist.
B.K.M. and J.M.W. conducted all of the experiments, helped in data analysis, and edited the manuscript. C.G.M. organized for sample collection and processing. J.K.N. assisted in study design and data interpretation. P.M.I. and O.N.N. organized study logistics, assisted in study design, and edited the manuscript. J.J. and A.L.R. provided assistance in study design, provided Rick17b assay, primers for MLST, and positive control DNA, and edited the manuscript. J.N.W. designed the study, directed the work, and drafted the manuscript. All authors have read and approved the final manuscript.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.