
Abstract
To clarify how Borrelia miyamotoi is maintained in the environment in Hokkaido, we examined Ixodes persulcatus for its prevalence among wild rodents and its tick vector by detecting a portion of the borrelial flaB gene in rodent urinary bladder and blood samples, and from whole ticks. We compared B. miyamotoi infection rates to Borrelia garinii and Borrelia afzelii, which are human Lyme disease pathogens also carried by wild rodents, and which are transmitted by the same vector tick. Whereas B. garinii and B. afzelii showed age dependence of infection rates among wild rodents (18.4% and 9.9% among adults and 6.0% and 3.4% among sub-adults, respectively) when looking at urinary bladder samples, B. miyamotoi infection rates were not age dependent for either blood (4.2% among adults, and 7.9% among sub-adults) or urinary bladder samples (1.0% among adults, and 1.7% among sub-adults). Moreover, while B. garinii and B. afzelii infection rates showed increases across months (June, July [p<0.05] and August [p<0.01] had higher rates than in May for adult rodents with B. garinii, and July and August had higher rates than in May [p<0.01] for adult rodents with B. afzelii), B. miyamotoi infection rates did not show significant month dependence. These differences in month and age dependence led us to suspect that B. miyamotoi may not develop persistent infections in wild rodents, as B. garinii and B. afzelii are thought to. Furthermore, we examined the extent of rodent exposure to I. persulcatus nymphs and larvae throughout most of the tick's active season (May through September), and determined that B. miyamotoi infection rates in sub-adult rodents were correlated with larval burden (p<0.01), suggesting that larvae may be very important in transmission of B. miyamotoi to wild rodents.
Introduction
Vector ticks have been shown to retain B. miyamotoi infections trans-stadially, and subsequent generations can be infected trans-ovarially; however, the infection rates have been noted to decrease between moltings, and not all larvae born from an infected female become infected (Scoles et al. 2001). Assuming that this is representative of B. miyamotoi infections of ticks in the environment, at least one reservoir species is necessary for maintenance of the spirochete. However, most studies on B. miyamotoi have focused strictly on its relationship with the vector tick, and few have tried to determine the reservoir species or evaluate the relationship of the pathogen with its reservoirs.
To determine how B. miyamotoi is maintained in an ecosystem, we examined wild rodents and questing I. persulcatus ticks for the presence of the borrelial flagellin gene (flaB), and also evaluated the exposure of rodents to I. persulcatus nymphs and larvae. Because rodents have been implicated as the most important reservoirs of B. garinii and B. afzelii, which are the pathogens of human Lyme disease in Japan (Nakao et al. 1994, Takano et al. 2011), and B. miyamotoi, which shares the same vector, we hypothesized that they share the same reservoirs. Therefore, we compared infection rates of B. miyamotoi among wild rodents trapped in Hokkaido, Japan, to the more studied B. garinii and B. afzelii.
Materials and Methods
Rodent trapping and tick collection
Two study sites were selected in Hokkaido, Japan, one at the base of the Shiretoko peninsula in eastern Hokkaido and the other near Shimizu, in central Hokkaido. Rodent trapping was performed using Sherman traps during 3- to 6-day stints in both locations each month from May through September of 2010 and 2011. Traps were initially placed at 5-meter intervals in two separate grids of 10×10 (200 traps), baited with oats, and were checked every 24 h. In July 2011, traps placed in central Hokkaido were reduced to 100 traps on a single grid, and traps placed in eastern Hokkaido were reduced to 40 traps on a single grid (4×10). For the remaining months of 2011 (August and September), only 40 traps on a single grid (4×10) were placed in each location. Trapped rodents were anesthetized with isoflurane in an open-drop system as described elsewhere (“Rodent anesthesia using open-drop exposure to isoflurane” 2010). Following cardiocentesis under anesthesia, rodents were promptly euthanized by cervical dislocation for sampling of the urinary bladder, which has been reported to develop persistent borrelial infections (Masuzawa et al. 1992). Feeding ticks attached to the neck and head were also removed and collected. All samples were stored at −20°C or −80°C until extraction of DNA. All rodent sampling was performed using methods approved by the Animal Care and Use Committee of Hokkaido University (Approval No. JU1105).
Between May and July of 2011, we collected questing nymph and adult ticks by flagging within vegetation with flannel cloth (approximately 1×1.5 meters) in and around the rodent trapping areas. Ticks were removed from the flags, and placed in BD Falcon™ conical tubes (BD Biosciences, San Jose, CA) with 21-gauge air holes along with several fronds of grass. Collected ticks were stored alive in the tubes sealed in plastic bags and refrigerated at 4°C for no more than 2 weeks or frozen at −80°C until DNA extraction.
Rodent and tick species identification and aging/staging
Rodents were identified to species based on morphologic characteristics (Abe 1994, Ohdachi et al. 2009). Individual rodents were also classified as adult (born prior to the sampling season) or sub-adult (born during the sampling season) based on tooth wear with a previously described 9 age-group classification scheme for Apodemus spp. (Hikida and Murakami 1980), and a previously described 6 age-group classification scheme for Myodes spp. (Abe 1976). Myodes spp. rodents were classified as adults if their teeth were designated as five or higher. In the case of Apodemus spp., because sub-adults born early in the season but trapped late in the season, often had tooth scores equivalent or higher than adults trapped in May, a graduated scale was implemented to remove early-season born rodents from late-season adult numbers. The lower-end cutoff for the adult designation was set at 4 and above in May, then consecutively raised one level each month until August (5 or higher for June, 6 or higher for July, and 7 or higher for August). Furthermore, because in September, sub-adults were often indistinguishable from adults, we classified all rodents trapped in September as sub-adults so that infection rates would be biased toward the sub-adult group rather than the adult group. All aging classification was performed by a single observer.
In 2011, feeding ticks collected from rodents were identified to species and stage based on described, morphologic differences (Yamaguti et al. 1971), and a tally was kept for each rodent. Questing ticks were also identified to species and stage using the same reference literature, and were then individually processed for DNA extraction.
DNA extraction, PCR, and sequencing
Urinary bladder DNA from mice was extracted using the Wizard® genomic DNA purification kit (Promega, Madison, WI) by the recommended protocol for soft tissue. DNA was extracted from clotted blood samples using a method described previously (Kanai et al. 1994). Questing tick DNA was extracted using the Wizard® genomic DNA purification kit with a modified protocol.
PCR was performed on DNA of rodent urinary bladders and blood, and on questing tick DNA for the presence of the borrelial flaB gene according to a previously described method that can detect as few as 10 spirochete cells in a sample (Sato et al. 1997, Takano et al. 2010). Confirmation of positive results was performed by replication. PCR products of positive samples were purified using ExoSAP-IT (GE Healthcare UK, Ltd., Buckinghamshire, UK) prior to direct sequencing on an ABI Prism 3130xl Genetic Analyzer (Life Technologies Co., Grand Island, NY). Sequencing results were compared to reference sequences recorded in GenBank for identification to species (B. garinii strain 20047, greater than 98% similarity, and B. afzelii strain VS461 and B. miyamotoi strain HT31, each greater than 99% similarity; accession numbers D86617, D63365, and D43777, respectively). Mixed sequencing results were resolved by TA cloning using either pGEM®-T Vector System I (Promega) with ECOS™ Competent E. coli (Nippon Gene Co., Ltd., Tokyo, Japan) or pCR™4-TOPO® with One Shot® TOP10 Chemically competent E. coli (Life Technologies Co.). Eight clones for each mixed sample were selected for re-sequencing and analysis. Once mixed samples were identified to species, they were counted as separate positives for each Borrelia sp. unless the mixture was of multiple strains of the same species, in which case, the sample was considered to be a single positive sample.
Results
Rodent species identification and aging
A total of 879 rodents were captured and included in this study. On the basis of aging criteria, 293 and 586 rodents were identified as adults and sub-adults, respectively. Five species were represented: Apodemus argenteus (n=137: adult=58, sub-adult=79), A. speciosus (n=446: adult=121, sub-adult=325), Myodes rex (n=12: adult=3, sub-adult=9), M. rufocanus (n=195: adult=62, sub-adult=133), and M. rutilus (n=89: adult=49, sub-adult=40).
Detection of borrelial DNA in rodent specimens
The urinary bladder of each of the 879 rodents was processed through to sequencing. Blood samples were processed and analyzed for 549 rodents, including 143 adults and 406 sub-adults (Table 1). A total of 139 urinary bladder samples (15.8%) and 40 blood samples (7.3%) were positive for one or more of the three borreliae of interest to this study (Table 1). B. garinii and B. afzelii were detected among 89 (10.1%) and 49 (5.6%) urinary bladder samples, respectively, whereas B. miyamotoi was rarely detected (1.5%). However, 38 of the 40 positive blood samples were identified as B. miyamotoi infections. Blood samples were available for 9 of the rodents positive for B. miyamotoi using the urinary bladder. All of these were also positive for B. miyamotoi (see supplementary material online at
B. afzelii and B. garinii mixed samples present (A. speciosus=9, M. rufocanus=1, M. rutilus=2, Total=12).
B. garinii and B. afzelii infection rates were both significantly higher for adults compared to sub-adults, whereas there was no significant difference between the two groups in the case of B. miyamotoi (Table 2). When calculated separately by species, A. speciosus was the only species showing a significantly higher infection rate of B. afzelli in adults when compared to sub-adults (adult=23/121, sub-adult=16/325, p<0.01, Fisher exact test). However, A. speciosus (adult=37/121, sub-adult=32/325) and M. rufocanus (adult=7/62, sub-adult=0/133) both had significantly higher (p<0.01, Fisher exact test) infection rates of B. garinii among adults. Although the overall infection rates and sample sizes were too small to determine significant difference for the other species, the general tendency was for infection rates of adults to be higher than sub-adults for both B. garinii and B. afzelii (data not shown). The exception was M. rex, for which no infections with either borrelia were detected. Given the above, only the adult and sub-adult totals including all species are reported (Table 2).
Significant difference in rates between age groups determined using Fisher's Exact test (P<0.01).
In contrast to B. garinii and B. afzelii, B. miyamotoi infection rates were not significantly different between adults and sub-adults for any of the rodent species for either sample type (i.e., for blood samples, A. speciosus: adult=3/62 [4.8%], sub-adult=21/229 [9.2%]; M. rufocanus: adult=2/22 [9.1%], sub-adult=8/84 [9.5%] [remaining data not shown]). Consequently, only the totals were reported for the adults and sub-adults of all species collectively for B. miyamotoi as well (Table 2). Furthermore, B. miyamotoi infection rates remained statistically stable for both adults and sub-adults and in total across all months, whereas both B. garinii and B. afzelii showed an increase in infection rates across months for adult and sub-adult groups with significantly higher values in later months (Fig. 1). Using the Kruskal–Wallis test with the Dunn multiple comparisons test, B. garinii infection rates among adults in June and July (p<0.05) and August (p<0.01) were all higher than in May. Among sub-adults, September had significantly higher infection rates than June (p<0.01) or August (p<0.05). B. afzelii infection rates were higher among adults in July and August than in May (p<0.01), and higher in July than in June (p<0.01). Sub-adults also showed an increase in B. afzelii with infection rates higher in September than in June, July, or August (p<0.01).
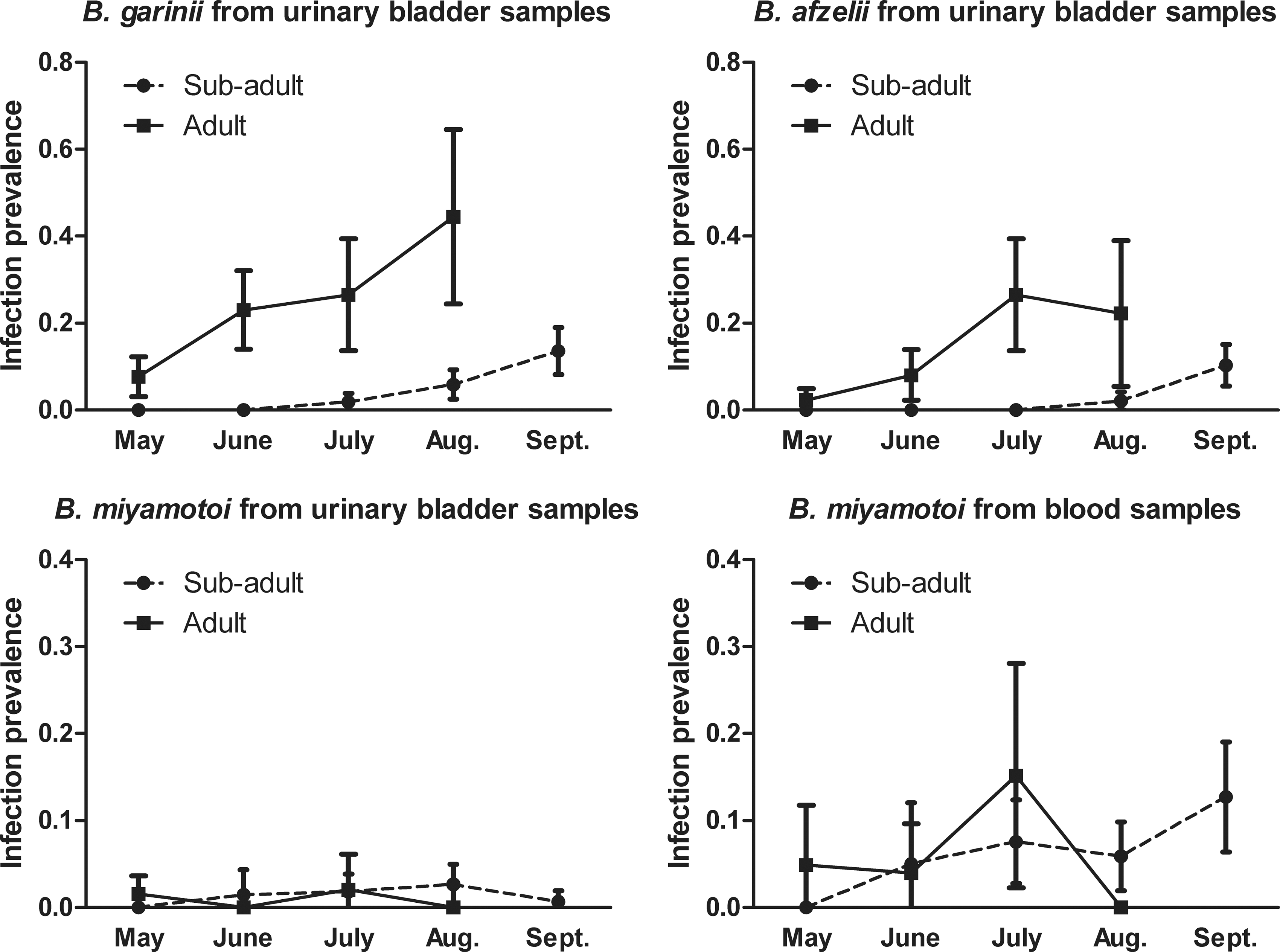
Borrelia-positive means by month with 95% confidence interval (CI) for urinary bladder and blood samples.
Feeding ticks collected from rodents
A total of 597 rodents trapped in 2011 and included in this study were examined for the attachment of feeding ticks. A total of 1604 ticks were found, and of those, 955 (59.5%) were identified as I. persulcatus larvae, and 177 (11.0%) were identified as I. persulcatus nymphs. In all, 275 rodents had I. persulcatus larvae attached and 80 had I. persulcatus nymphs attached (see supplementary material online at
Using the Mann–Whitney test, there was no significant difference between the mean number of I. persulcatus larvae attached to adults and sub-adults of all rodent species collectively (adults, mean=1.5, standard deviation [SD]=2.7, n=199; sub-adults, mean=1.6, SD=3.2, n=398). Upon analysis of total adults and sub-adults for the mean number of nymphs attached, however, the difference was significant (p<0.01), with adults accounting for a larger portion of the nymphs (adults, mean=0.6, SD=1.5, n=199; sub-adults, mean=0.1, SD=0.6, n=398).
Tick burdens across months were charted for both I. persulcatus larvae and nymphs (Fig. 2). Rodents trapped in August had a significantly lower mean burden of larvae than all other months (p<0.01, Kruskal–Wallis with the Dunn multiple comparisons test). Also, the mean larval burden of rodents trapped in September was significantly higher than that of rodents trapped in July (p<0.01, Kruskal–Wallis with Dunn). As for the mean burden of nymphs, rodents trapped in June had a significantly higher burden than those trapped in August or September (p<0.01 and p<0.05, respectively, Kruskal–Wallis with Dunn).

(Top graphs) Counts (with means and 95% confidence interval [CI]) of nymphs and larvae for rodents which had one or more nymphs attached. (Bottom graphs) Mean number of nymphs and larvae attached per rodent in total (with 95% CI).
Cross-evaluation, using the Fisher exact test, of positive results for borreliae and the number of I. persulcatus larvae or nymphs attached to rodents showed that B. garinii positive, adult rodents were more likely to have both nymphs and larvae attached (p<0.01 and p<0.05, respectively) than negative adults. Sub-adult rodents showed no such association for B. garinii. Sub-adult rodents that were positive for B. miyamotoi, on the other hand, were more likely to carry a larval burden than negative rodents when considering results from either urinary bladder or blood samples (p<0.05 and p<0.01, respectively). Adult rodents positive for B. miyamotoi showed no correlation with either nymph or larval numbers. B. afzelii-positive rodents showed no significant association with larval or nymph numbers.
Questing ticks
Of a total 240 I. persulcatus ticks collected by flagging and included in this study, 163 were adults, and 77 were nymphs. PCR and sequencing analysis indicated that 49 (30.1%) adults and 6 (7.8%) nymphs were positive for the borrelial flaB gene. Among the adult samples, 44 (27.0%) were positive for B. garinii, 5 (3.1%) were positive for B. afzelii, and 3 (1.8%) were positive for B. miyamotoi. Of the nymphs, 6 (7.8%) were positive for B. garinii, and 1 (1.3%) sample was positive for B. afzelii. None of the nymphs we tested carried B. miyamotoi.
Discussion
In the present study, we attempted to elucidate the ecology of the borrelial spirochete B. miyamotoi, a potential human pathogen in Japan. Although it is well documented that B. miyamotoi is carried by ixodid ticks, the natural reservoirs have not yet been determined. We provide relative infection rates of multiple rodent species found in Hokkaido, Japan, and also show that the ecology of B. miyamotoi within wild rodents differs from that of B. garinii and B. afzelii, which are also transmitted by I. persulcatus ticks.
Our results, showing the highest B. garinii and B. afzelii infection rates among A. speciosus, are consistent with a previous study in Hokkaido (Nakao et al. 1993), suggesting that sampling and testing methods were appropriate. For B. miyamotoi, however, significant differences could not be determined among species with the current sample sizes, although trends suggest that A. speciosus and M. rufocanus may harbor higher infection rates than the other species. This may be important, given that these are prominent rodent species in Hokkaido (Forestry Research Institute 2011). Regardless, because no statistical difference was found, we grouped all species together to compare the difference in infection rates of the borreliae between adult and sub-adult rodents.
We interpret the significantly higher infection rates of B. garinii and B. afzelii among adults compared to sub-adults, as consistent with persistent infection, which has been shown to occur for Borrelia burgdorferi, another Lyme disease pathogen, in rodents (Schwan et al. 1991). Whereas B. garinii and B. afzelii showed this age dependence, B. miyamotoi did not. The lack of age dependence for B. miyamotoi infection, along with the absence of significant difference in infection rates of the bladder across months, suggests that, unlike B. garinii and B. afzelii, it does not result in persistent infections of the urinary bladder. It has also been shown that skin samples from mice had lower infection rates than blood (Barbour et al. 2009), and evaluation of spleen samples suggested that this organ is also not a focus of persistent infection (data not shown). However, we cannot yet exclude the possibility that B. miyamotoi persists in other organs. B. duttonii, another tick-borne relapsing fever Borrelia sp., for instance, is reported to remain in the brain of mice in the absence of bacteremia, and recurrence of bacteremia can be induced by stress (Larsson et al. 2006). Furthermore, we have not evaluated the possibility that rodents infected with B. miyamotoi have reduced life spans.
Given the data that age is not positively correlated with B. miyamotoi infection, we propose that B. miyamotoi infection rates are largely due to I. persulcatus larval burden. Using both urinary bladder and blood samples, sub-adult rodents positive for B. miyamotoi were more likely to carry larvae. Given this, the fact that larvae are responsible for a far larger portion of the overall tick burden than are nymphs, and the fact that larvae are known to harbor B. miyamotoi infections in the United States (Richter et al. 2012), it seems reasonable that the majority of transmission events from tick to rodent be due to larvae (whether vertical transmission of B. miyamotoi occurs among ticks in Japan still needs to be determined). This is in direct contrast to B. garinii and B. afzelii, for which vertical transmission is thought to seldom occur, and therefore cannot be transmitted to rodents by larvae. The lack of correlation between B. miyamotoi-positive adults and larval burden may simply be due to the small number of samples.
Although statistically insignificant, B. miyamotoi infection rates in blood show a mild incline suggestive of month dependence. However, because B. miyamotoi was deemed not age dependent, we suspect this incline reflects trends in I. persulcatus larval burden. The two graphs are suggestive of parallel change with a mild lag of infection compared to larval burden, as we would expect.
The 1.8% prevalence of B. miyamotoi among questing adult I. persulcatus ticks from our study areas was similar to the infection rates found among ticks of the same species in sampled areas of Russia and among I. scapularis ticks in parts of the northeastern United States (Scoles et al. 2001, Fomenko et al. 2010). The consistently low prevalence of B. miyamotoi among ticks and rodents compared to prevalences of B. garinii and B. afzelii may be, in part, associated with the unique ecology of the spirochete (e.g., the possible absence of persistent infections).
In conclusion, we suspect that, unlike B. garinii and B. afzelii, which cause persistent infections in the urinary bladder, B. miyamotoi may not. Furthermore, we predict that the prevalence of B. miyamotoi among wild rodents is largely a consequence of transmission from I. persulcatus larvae, and that infection rates among rodents may reflect seasonal changes in the population of questing larvae. However, further research, including infection trials, is necessary to fully substantiate these hypotheses.
Footnotes
Acknowledgments
The authors thank Mr. Hidenori Nishizawa for his help in sampling, Drs. Satoshi Ohdachi and Jiro Arikawa for use of their traps, Dr. Hisashi Abe for instruction on rodent identification and aging, and Ms. Kozue Sato and Ms. Chieko Sugimori of the National Institute of Infectious Diseases for their help in cloning and related DNA work. This research was funded in part by the Mitsui & Co., Ltd. Environment Fund, in part by a grant for research on emerging and reemerging infectious diseases from the Japan Ministry of Health, Labor, and Welfare, and by the Global COE Program “Establishment of International Collaboration Centers for Zoonosis Control,” MEXT, Japan.
Author Disclosure Statement
This paper was written to partially fulfill the requirements for a Ph.D. degree at the Hokkaido University, Graduate School of Veterinary Medicine, for the lead author, Kyle Taylor. No other authors have any interests that conflict with the publication of this paper.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.