
Abstract
The aim of this study was to analyze synanthropic birds as risk factors for introducing ticks and tick-borne pathogens into human settlements, with an emphasis on rickettsiae. Altogether 184 subadult ticks were found on 5846 birds. Tick infestation was most prevalent during the spring. In this sample group the majority of ticks were molecularly identified as Ixodes ricinus, and three individuals collected from the European robin as Hyalomma marginatum marginatum. The latter is the first molecularly confirmed occurrence of this species in Hungary. Rickettsia aeschlimannii was detected in H. marginatum, also for the first time in Hungary, and in ticks from an urbanized bird species north of the Mediterranean countries. The overall prevalence range of rickettsiae (including R. helvetica and R. monacensis) in ticks of synanthropic birds was 29–40%, exceeding that in questing ticks of relevant species reported earlier. Additionally, in specimens of I. ricinus, the presence of Borrelia burgdorferi sensu lato (s.l.), Anaplasma phagocytophilum, and a new Francisella-like genotype was also verified. Thus, it can be concluded that birds with urban or periurban habitats pose a high risk as tick carriers and reservoirs of zoonotic agents, especially of rickettsiae.
Introduction
The significance of birds in these situations involving zoonotic pathogens may be particularly high in urban and periurban habitats, where, having flown in, their engorged ticks can detach. Thus, inflying birds may promote shorter-term changes than already present local reservoirs. Irrespective of this, most of the studies on ornithophagous ticks and associated pathogens focus on the way birds most likely have become tick infested, i.e., during ground feeding (Elfving et al. 2010, Dubska et al. 2011). Several reports concern exotic and anthropophilic ticks or zoonotic agents that may be imported by migratory birds to nonendemic regions, but they do not approach avian hosts according to their different potentials for arriving in a human environment (Rumer et al. 2011a, Jameson et al. 2012). To provide relevant information on the latter aspect, it is inevitable to address the higher or lower impact of tick dispersal by birds in the context of their preferred habitats.
Therefore, this study was undertaken not only to collect and molecularly analyze ticks from birds, but also to highlight the epidemiological role of those species that have urban/periurban habitats and to compare the prevalence rates of bacterial agents in ticks of synanthropic birds with those in other geographically relevant (Hungarian, European) sample sources. Such data on borreliae are available in the literature (Dubska et al. 2011), but are lacking for rickettsiae.
Materials and Methods
Sample collection and preparation
Birds mist-netted at the Ócsa Ringing Station (Duna-Ipoly National Park) were checked for the presence of hard ticks (Acari: Ixodidae) during 2011. Specimens removed with forceps were put into 70% ethanol. Subadult ticks were morphologically identified by using standard keys (Babos 1964): Haemaphysalis (Ha.) concinna individuals on the species level, whereas Ixodes and Hyalomma (Hy.) specimens were identified only on the genus level. In the latter two genera, species were determined molecularly because of the morphological similarity of immature stages of different species to each other (Rumer et al. 2011a, b). Focusing on migratory birds, spring-collected samples were selected for further analysis. To minimize sample number for multiple analyses, ticks were processed individually or in pools (of 2–7 specimens) separated according to their host individuals, species, and developmental stages. DNA was extracted as described (Hornok et al. 2010a).
Molecular identification of subadult ticks
In case of Hyalomma subadults, fragments of the 16S and 12S rRNA genes were amplified in two PCR assays, as already reported (Black and Piesman 1994, Norris et al. 1999). PCR products were resin purified (Wizard, Promega) and cloned into the pGEM-T vector (Promega, Madison, WI) for sequencing both strands (Secugen S.L., Madrid, Spain). At least 3 clones were sequenced for each amplicon. Ixodes subadults were identified with 2 species-specific primer sets, 1 for I. ricinus (dITS678, rITS814) and another for I. persulcatus (dITS118, rITS436) according to Rumer et al. (2011b).
Molecular analyses for tick-borne agents
PCR assays for pathogen detection in tick DNA are summarized and referenced in Table 1. These tests were performed with all individual and pooled samples. For quantitation by standards in the Rickettsia helvetica 23S rRNA TaqMan PCR, 10-fold serial dilutions of cloned plasmid DNA with known copy numbers were used. From Rickettsia gltA TaqMan PCR-positive samples with threshold cycle (Ct) values ≤30, a 381-bp portion of the citrate synthase gene was sequenced as described (Boretti et al. 2009). Francisella-like bacteria were analyzed as reported (Barns et al. 2005), and phylogenetic comparison was made by using the neighbor-joining method with maximum composite likelihood model and 1000 bootstraps with the MEGA5 program (Tamura et al. 2011).
Statistical methods
Prevalences were compared by the Fisher exact test, and differences were regarded significant if p ≤0.05. Prevalence rates were calculated from PCR results of individual samples. For pooled ticks, the minimum prevalence was determined from the number of positive pools, expressed as the percentage of all evaluated tick individuals of the same species (provided that there must have been at least one PCR-positive specimen in each PCR-positive pool). Overall prevalence range of a pathogen group is given as minimum prevalence to prevalence based on PCR results of all individual samples.
Results
Species and abundance of ticks
Altogether 184 subadult ticks were found on 5846 birds. Tick infestation was significantly more prevalent during the spring (March–May, 62 of 1786 birds), than in the summer (June–August, 17 of 2630 birds) or autumn (September–November, 16 of 1430 birds) (p<0.001). In the spring and in the autumn, the majority of ticks were Ixodes subadults (96.3% and 100% of all collected specimens, respectively), whereas in the summer highest abundance (92%) was observed in case of Ha. concinna larvae and nymphs.
Mean tick numbers per host (intensities of infestations) in the spring (i.e., during migration) were higher among bird species known to have urban/periurban habitats, than in the case of nonsynanthropic ones, whereas the prevalence rates of tick infestations were more equilibrated between these groups (Table 2).
Recaptures are not included. Except for Acrocephalus spp. and Locustella luscinioides, all bird species in this table are considered to have urban or periurban habitats. Migration types of bird species are also indicated (i.e., N=nonmigrant, S-MD short to middle distance and LD=long distance migrant).
L, Larva; N, nymph.
Prostriate subadults were molecularly identified as I. ricinus. Three Hyalomma immatures (2 molting larvae and 1 nymph) were also collected from a highly urbanized species, the European robin (Table 2). Molecular analysis of these revealed 100% sequence homology to Hy. marginatum marginatum from Morocco (accession numbers L34307 and AF150034 for 16S and 12S rRNA genes, respectively).
Species and prevalence of tick-borne agents
In the spring, 108 ticks were removed from birds. These were analyzed as 56 individual samples and 16 pools. Half of them (36 of 72) were PCR positive for at least 1 of the tested pathogens (Table 3). Except for 1 I. ricinus nymph, dual infections were found in pooled samples.
Except for Locustella luscinioides, all bird species in this table are considered to have urban or periurban habitats. All samples were evaluated for all pathogens, but only the positives are shown numerically. Samples with simultaneous presence of R. helvetica and B. burgdorferi sensu lato (s.l.) are marked with asterisk.
In the R. helvetica-specific PCR, 17 samples were positive (3/16 of I. ricinus larvae, 13/50 of I. ricinus nymphs, 1/3 of H. concinna nymphs; Table 3), meaning 15.7% minimum prevalence. Copy numbers per 5 μL were 1–102 in 8 samples, 102–104 in 1 sample, and higher (maximum 2.8×105) in a further 8 samples. Regarding other rickettsiae, 15 samples were positive (2/16 of I. ricinus larvae, 10/50 of I. ricinus nymphs, 3/3 Hyalomma subadults; Table 3). Sequencing from 10 samples of I. ricinus showed 100% homology to a former Hungarian isolate of Rickettsia monacensis (accession number EU853830). The minimum prevalence for this species was 11.1%. In Hy. marginatum, Rickettsia aeschlimannii was identified, for the first time in Hungary, with 100% sequence similarity to an isolate from Egypt (HQ335153). The overall prevalence range for rickettsiae in ticks of synanthropic birds was 29–40%.
PCR positivity to Borrelia burgdorferi sensu lato (s.l.) was detected only in I. ricinus (1/16 samples of larvae and 7/50 of nymphs were positive; Table 3), amounting to a minimum prevalence of 7.4% (Table 3). Results of this assay (low copy numbers, Ct >30) did not allow genotype sequencing. In the dually positive 4 samples, borreliae were always in concomitancy with R. helvetica (Table 3).
Anaplasma phagocytophilum was demonstrated in 1 pool of I. ricinus nymph from Erithacus rubecula, with simultaneous positivity to R. monacensis. Interestingly, from 1 I. ricinus larva, also from E. rubecula, a novel Francisella-like sequence was obtained (1036 bp, accession number JQ740890), which showed the closest (99%) similarity to endosymbionts detected in Dermacentor reticulatus in central Europe (Fig. 1). All samples were negative for Coxiella burnetii.
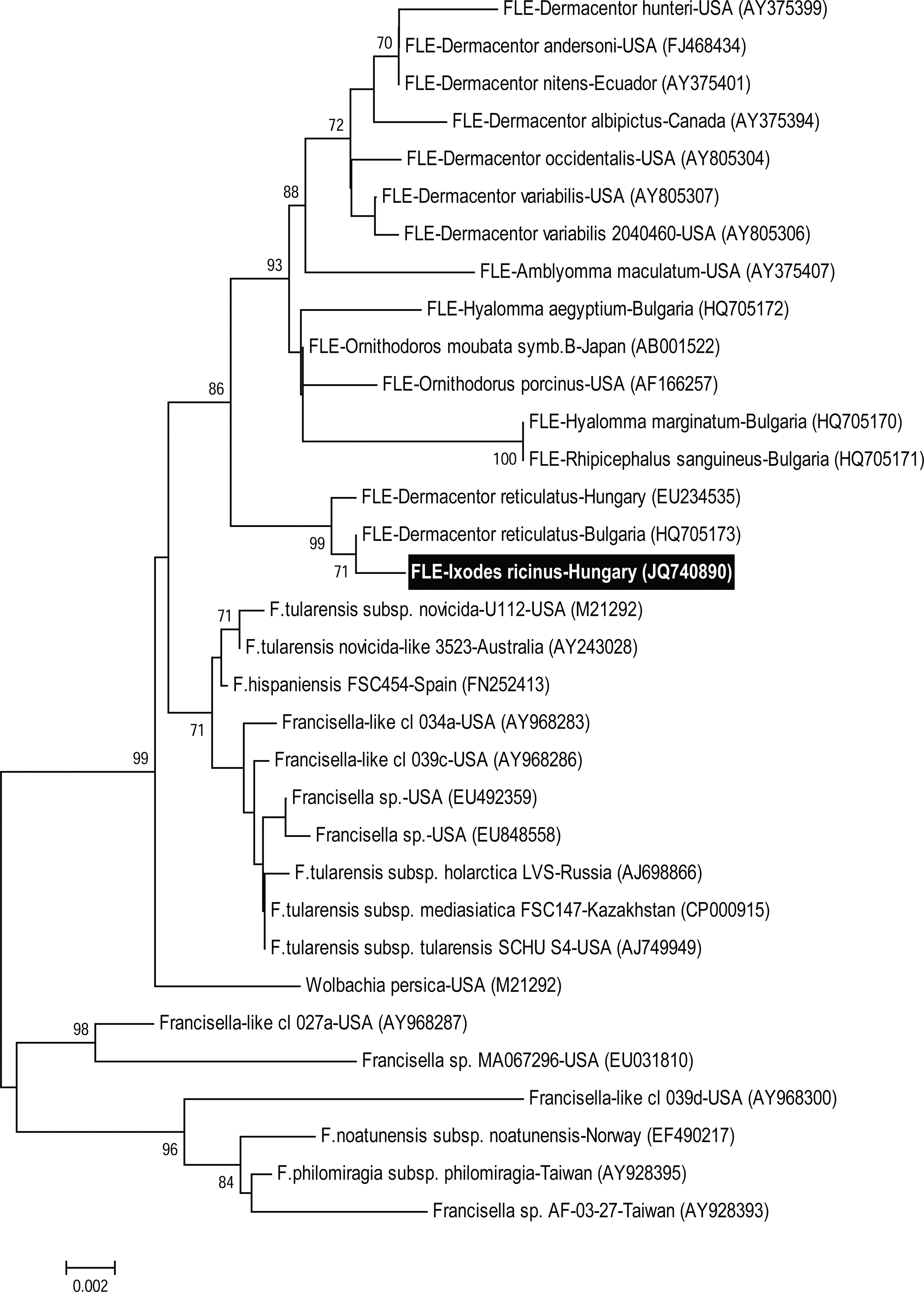
Neighbor-joining phylogenetic tree showing relationships of 16S rRNA gene sequences obtained from Francisella species and Francisella-like endosymbionts (FLEs) with the novel Francisella-like isolate from Ixodes ricinus larva. Bootstrap values of neighbor-joining (1000 replicates) of ≥70 are shown.
Discussion
This is the first survey focusing on the potential role of synanthropic birds in spreading tick-borne rickettsiae. Tick samples collected from avian hosts during the spring also allowed verification for the first time with molecular methods that nonendemic tick and Rickettsia spp. are imported by migratory birds to central Europe. The most important species in this respect, the European robin (E. rubecula), arrives in Hungary from the Mediterranean countries (Csörgő et al. 2009). Similary, the majority of bird species evaluated in the present study are short- to middle distance migrants (Table 2), most frequently caught at Ócsa Ringing Station on their way to northern Europe (Csörgő et al. 2009). Because birds can cover few hundreds of kilometers during a single day of migration, and tick larvae and nymphs are known to attach and feed for several days (Babos 1964), we can assume that most of the ticks and tick-borne pathogens identified in the present study from the spring collection derived from southern Europe.
Taking into account that the majority of ixodid ticks in Hungary and surrounding countries have spring peak activities (Babos 1964, Hornok 2009), it was not unusual to find the highest prevalence of tick infestation among birds in that season, although this is in contrast to some other observations in the region (Dubska et al. 2011). Summer dominance of Ha. concinna in this study also reflected the pattern already reported for questing ticks in the country (Hornok 2009).
Hyalomma spp. are rarely found in Hungary. Their subadults were either not identified on the species level (Janisch 1962), or were most likely members of the Hy. marginatum complex, but were named synonymously as Hy. impressum (Babos 1964, Apanaskevich and Horak 2008). One Hy. marginatum nymph was also morphologically identified in a recent study (Földvári et al. 2011). Correspondingly, while the sporadic import of Hy. marginatum subadults has been long accepted in Hungary, here we report the first molecular evidence of this species in the country.
In Hy. marginatum subadults of the present study, R. aeschlimannii was identified for the first time in Hungary. This human pathogenic rickettsia is endemic to the Mediterranean part of Europe (Fernández-Soto et al. 2003, Santoz-Silva et al. 2006) and was recently detected in the same tick species from a migratory bird in Germany (Rumer et al. 2011a). However, the sample source in that study was the reed warbler (Acrocephalus scirpaceus), which is a nonurban bird species unlikely to disperse R. aeschlimannii-infected ticks in a human environment. On the other hand, here R. aeschlimannii was demonstrated in ticks of the European robin known for its synanthropic habitats. Therefore, the present data are the first to indicate that R. aeschlimannii can be imported by migratory birds into human settlements north of the Mediterranean basin.
The prevalence range of Rickettsia spp. in ticks of synanthropic birds in the present study was 29–40%. This means that birds carried ticks, predominantly I. ricinus, with a higher prevalence of rickettsiae than shown before with the same molecular methods in the case of questing ticks in the region (minimum prevalence of R. helvetica in Ha. concinna, I. ricinus was 1–7%; pool prevalence of R. monacensis was 16%: Hornok et al. 2010b). In another study using similarly sensitive real-time PCR on samples from northern Europe, but regardless of synanthropic avian hosts, the prevalence of rickettsiae in bird ticks was only 11% (Elfving et al. 2010). In addition, the prevalence of R. aeschlimannii in Hy. marginatum in foreign studies was significantly (p<0.001) lower, 6–9% (Fernández-Soto et al. 2003, Santoz-Silva et al. 2006). Thus, these data indicate a high risk for import of tick-borne rickettsiae by birds into urban/periurban habitats. One possible explanation for higher prevalence of rickettsiae in engorged ticks is that their hosts were rickettsemic. Alternatively, the distribution of these zoonotic agents may be biased in ticks of the human environment, and in this way more infected ticks can attach to birds with this type of habitat. Data in the literature support the urban/periurban accumulation of tick-borne rickettsiae, although this may depend on the species (Raoult et al. 1987, Milagres et al. 2010). R. monacensis, which had a high prevalence in ticks of synanthropic birds in this study, was also isolated first in a city park (Simser et al. 2002).
Higher prevalence of B. burgdorferi s.l. in nymphs, rather than in larvae of I. ricinus, here corresponds to the low frequency of transovarial transmission of borreliae (Hubálek 2004). Because bird-feeding ticks in the present study had a higher prevalence of both B. burgdorferi s.l. and A. phagocytophilum than ticks collected from rodents in the same region (Rigó et al. 2011), these data are in line with reports on the higher epidemiological significance of birds as hosts of ticks and as reservoirs of these tick-borne pathogens in comparison with small mammals (Franke et al. 2010).
Despite the fact that C. burnetii was recently shown to be highly prevalent in samples of domestic ruminants in Hungary (Gyuranecz et al. 2012), and birds are long known carriers of this agent (Babudieri and Moscovici 1952), here all ticks were found to be PCR negative. This might mean that birds and/or their ticks do not play an important role in the transmission of Q fever in the region and other reservoirs participate in its sylvatic cycle.
One Francisella-like sequence was also obtained from an I. ricinus larva. The relevant tick most likely acquired this bacterium transovarially, because an engorged nymph from the same bird was PCR negative. However, the horizontal transfer, e.g., by co-feeding, from other ticks can not be excluded, as suggested in case of the ancestors of these endosymbionts (Scoles 2004). Francisella-like agents were only detected so far in D. reticulatus and Hyalomma sp. ticks (Escudero et al. 2008, Sréter-Lancz et al. 2009, de Carvalho et al. 2011, Ivanov et al. 2011); therefore, this is the first molecular evidence of their occurrence in I. ricinus. Francisella-like bacteria were also reported from small mammals, and they may even have some degree of pathogenicity in these hosts (Escudero et al. 2008). Here it is indicated for the first time that they may associate with bird ticks. The sequence divergence between the Francisella-like agent in the present study and those already reported from D. reticulatus exceeds in magnitude the difference between Francisella tularensis ssp. tularensis and F. t. ssp. holarctica in the same part of their 16S rRNA genes.
In summary, the epidemiological significance of migratory birds importing exotic tick species and arriving in gardens and city parks around springtime may be considerably high. Synanthropic birds are also important to take into account during tick-borne disease risk assessment, because ticks disseminated by them may increase the local prevalence of rickettsiae and sometimes may introduce even nonendemic pathogens.
Footnotes
Acknowledgments
S. Hornok was supported by the Bolyai János scholarship (Hungarian Academy of Sciences).
Author Disclosure Statement
No competing financial interests exist.