
Abstract
Despite its confirmed establishment in neighboring Greece and Bulgaria, the presence of the Oriental invasive species Stegomyia albopicta (Skuse) (=Aedes albopictus) has never been confirmed in Turkey. Active surveillance for this container-breeding species was carried out using oviposition traps at 15 discrete sites in the towns of Ipsala (n=8 sites), Kesan (n=5) (Edirne District), and Malkara (n=2) (Tekirdag District) in the Thrace region of northwestern Turkey, from May 23 through November 10, 2011. Eggs collected were reared to the fourth larval instar and adult stages where possible to facilitate integrated morphological and molecular species identification. DNA barcodes (658 bp of the mitochondrial cytochrome c oxidase I [COI] gene) were compared with all four potentially invasive Stegomyia species: St. aegypti, St. albopicta, St. cretina, and St. japonica. Sequences generated for samples collected in Thrace Region were herein confirmed as St. albopicta, the first record of this vector species in Turkey. Eggs of St. albopicta were detected in two discrete localities: (1) In the grounds of a restaurant in Kesan (in week 36), and (2) in the customs area of the Turkish–Greek border at Ipsala (in weeks 32 and 38). Multiple detection of St. albopicta eggs indicates the possible establishment of the species in northwestern Turkey. Finding this important disease vector has implications for public health and requires the implementation of active vector monitoring programs and targeted vector suppression strategies to limit the spread of this invasive vector species in Turkey.
Introduction
Under laboratory conditions, St. albopicta is shown to be able to function as a bridge vector for West Nile virus (WNV) between the enzootic Culex spp.–avian cycle and susceptible mammalian hosts, including humans (Turell et al. 2001, Sardelis et al. 2002). St. albopicta is also implicated in zoonotic disease cycles, indicating that the species also feeds on other mammalian hosts. St. albopicta is a competent vector of the filarial nematodes Dirofilaria immitis and D. repens and is a natural vector of D. immitis in Italy (Cancrini et al. 2003a,b). Because of its aggressive anthropophilic behavior, St. albopicta could also enhance the zoonotic circulation of filarial nematodes from animals to humans in an urban environment (Cancrini et al. 2003a).
The global invasion of St. albopicta, originally native to subtropical southeastern Asia, islands of the Western Pacific and Indian Ocean, into the Pacific islands, Africa, the Caribbean, the Middle East, the Americas, and Europe, has been well documented (e.g., Gratz 2004, Benedict et al. 2007). The species was first detected in Europe from Albania in 1979 (Adhami and Murati 1987), and its establishment has been reported in 12 further European countries and microstates to date: Croatia, France, Greece, Monaco, Montenegro, Italy, Malta, San Marino, Slovenia, Spain, Switzerland, and Vatican City (Benedict et al. 2007, Scholte et al. 2007, ECDC 2009, Gatt et al. 2009). The presence of St. albopicta in Belgium, Bosnia and Herzegovina, Germany, and The Netherlands has been documented, but these reports are believed to represent accidental importations, rather than established populations at present (Scholte et al. 2007, ECDC 2009, Gatt et al. 2009). In addition, the presence of St. albopicta in Bulgaria was reported at a scientific meeting in 2011, but as yet this remains unpublished (Ognyan Mikov, pers. comm.); the species has been recently identified in Bolshoi Sochi Region of Russia (Anonymous 2012).
Distribution models predict that St. albopicta will continue to expand depending on transport, environmental, and climatic changes (Knudsen et al. 1996, Medlock et al. 2006, Benedict et al. 2007). The majority of St. albopicta introductions worldwide are reportedly due to dormant egg transfer via the international trade in used tires (Reiter and Sprenger 1987) and shipments of the Asian plant “lucky bamboo” (Dracaena spp.) (Linthicum et al. 2003, Scholte et al. 2007) or by the inadvertent transport of adult mosquitoes in aircraft and other modes of transport (Gratz et al. 2000). Between 1989 and 2010, Turkey imported 1627 tons of used tires from 20 countries, including United States, Greece, Japan, South Korea, China, Taiwan, Thailand, and Singapore, where St. albopicta is known to be established and/or endemic (Unlu and Farajollahi 2012).
Turkey has been identified as one of several countries (Cyprus, Bulgaria, Macedonia, Portugal, and southern Russia) under high risk of invasion due to its favorable climatic conditions and geographical proximity to established populations of St. albopicta (Mitchell 1995b, Knudsen et al. 1996, Benedict et al. 2007, ECDC 2009, Fischer et al. 2011, Caminade et al. 2012, Unlu and Farajollahi 2012). The suitability for the establishment of St. albopicta in Turkey has been confirmed by risk maps using different analytical techniques (e.g., Random Forest model, Multi-Criteria Decision Analysis, 0 and −5°C isotherm analysis, mean yearly temperatures, annual rainfall) and shows highest suitability in the coastal parts and lower likelihood of establishment in mountainous regions (ECDC 2009, Caglar and Karacaoglu 2011, Unlu and Farajollahi 2012).
Turkey has 18 major ports and 11 border gates, which are mostly located in areas with favorable climatic conditions for St. aegypti establishment. The Thrace region embraces both the Kapikule (Turkey–Bulgaria) and Ipsala (Turkey–Greece) border gates, which are the main trade routes between Turkey and the rest of Europe. This part of Turkey was considered at especially high risk, because St. albopicta is already established in northern regions of neighboring Greece (Corfu, Igoumenitsa, and Serres) (Samanidou-Voyadjoglou et al. 2005, Unlu and Farajollahi 2012). Following the advice of the European Centre for Disease Prevention and Control (ECDC) (2009), an active surveillance program was established in the Thrace Region of northwestern Turkey near to the Greek border in the summer of 2011, with the explicit aim of determining whether this invasive vector species was present and/or established in Turkey.
Materials and Methods
Study area
Fifteen sites in three districts (Ipsala, Kesan, and Malkara) were selected for active surveillance in the Thrace Region of northwestern Turkey, based on the climatic suitability of the region for St. albopicta establishment as stated by previously determined risk maps (ECDC 2009, Unlu and Farajollahi 2012) (Table 1, Fig. 1).
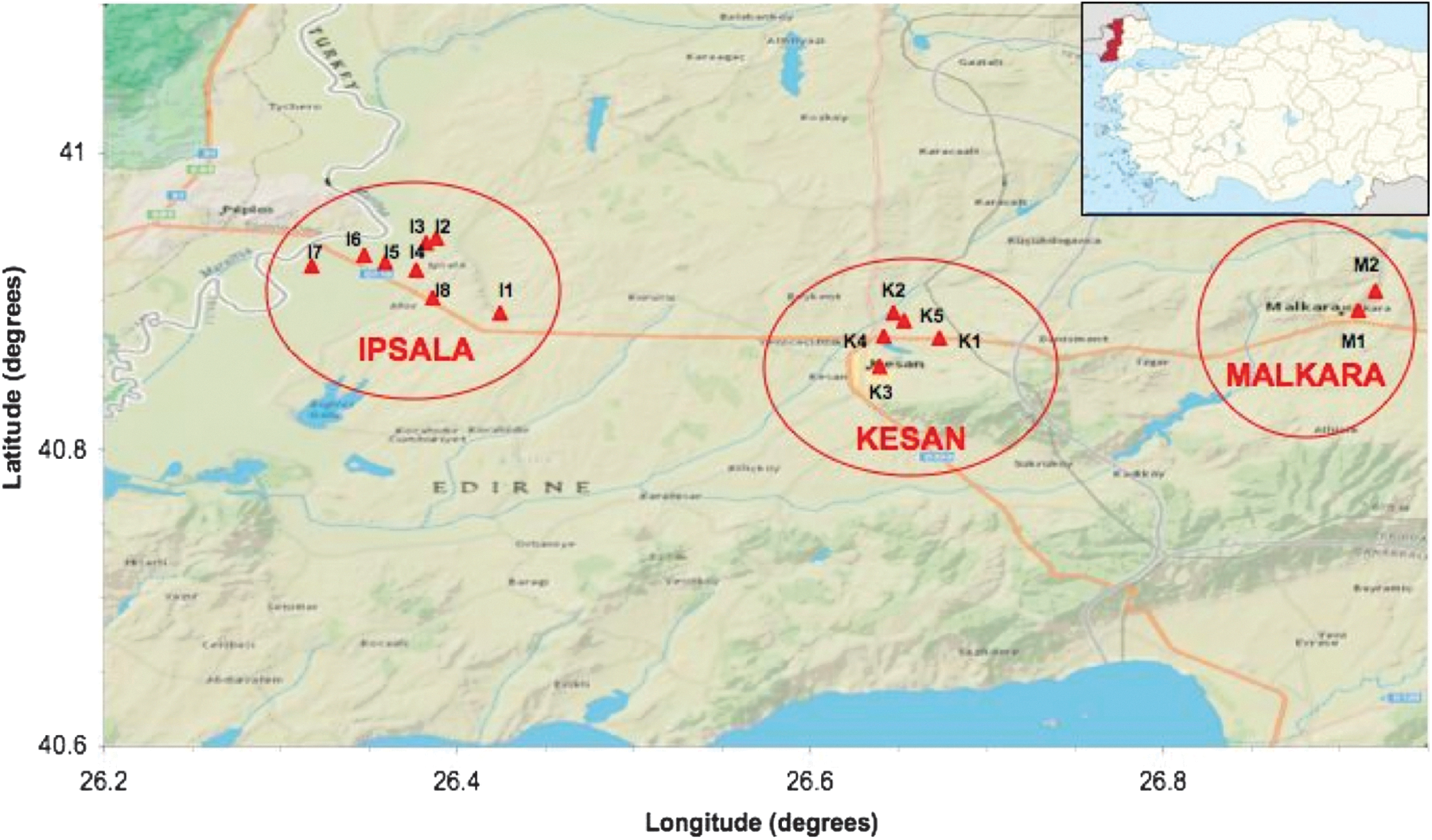
Map detailing the localities of sampling stations in the Thrace Region of northwestern Turkey. I: Ipsala, (1) I1: Truck parking area, (2) I2: Buket Street, (3) I3: Cikmaz Street, (4) I4: Building gardens, (5) I5: Government fish breeding farm, (6) I6: Ipsala customs area, (7) I7: Rice cultivation company, (8) I8: Tire repair shop. K: Kesan, (9) K1: Truck parking area, (10) K2: Supermarket parking area, (11) K3: University campus, (12) K4: Park, (13) K5: Restaurant gardens. M: Malkara, (14) M1: Cami Street, (15) M2: Exhibition Center gardens (see Table 1 for site coordinates).
K, Kesan District; I, Ipsala District; M, Malkara District.
Ipsala province (106 km2; population 8033) is located 108 km southwest of Edirne city and 2 km east of the Greek border. The customs gate in this area represents the most intensive transport route between Greece and Turkey for import/export of goods, and facilitates high numbers of person transits between the nations, including tourists. The north and eastern parts of Ipsala are surrounded by small hills (100–300 meters), whereas the Ipsala plain, with its intensive rice cultivation, dominates the western area. The natural borders of Ipsala correspond to the Meric River in the west and the Ergene River in the northwest. Continental climate prevails in Ipsala, with considerable seasonal and daily fluctuations in temperature. Intensive wet rice cultivation increases the relative humidity in summer (Ipsala Government 2012).
Kesan province (1087 km2; population 54,367) is 112 km south of Edirne, some 30 km east of Ipsala province and 26 km west of Malkara province, respectively. The province is dominated by lowland plains (∼100 meters above sea level [a.s.l]), but the Korudag mountains in the southeast rise to 371 meters a.s.l. The Marmara type of Mediterranean climate prevails in Kesan, which is more moderate at the southern coastal areas on the Aegean Sea (Saros Bay). The mean annual precipitation is 48.8 mm with 75.6% relative humidity, and the mean annual temperature is 13.7°C in Edirne. During the mosquito season, from May to September, the mean relative humidity is 72% and mean temperature is 21.8°C (min 11.4°C and max 31.7°C) (Kesan Government 2012).
Malkara Province (1225 km2; population 28,788) lies 56 km west of Tekirdag and shares a similar geography to Kesan, composed mainly of lowland plains. The Tekir Mountains, 25 km south of Malkara, have their maximum altitude at Ganos temple (845 meters a.s.l.). A semicontinental climate prevails in Malkara, which is a transition type between the Mediterranean and Black Sea climates. The mean annual precipitation is 48.3 mm with 85% relative humidity. During the mosquito season, from May to September, the mean relative humidity is 79% and mean temperature is 21°C (minimum 12.4°C, maximum 27.9°C) (Malkara Government 2012).
Surveillance program
Surveillance activities were conducted between May 23 and November 10 (weeks 22–42) of 2011. Guided by the risk analysis reports (ECDC 2009, Unlu and Farajollahi 2012), we considered the main possibility of entry to be the passive transportation of adults/eggs from Greece via the main trade motorway (E84). Therefore sampling stations were determined at urban and periurban areas along the E84 motorway (Fig. 1). Some 54 ovitraps were set up at 15 locations (Ipsala, n=8; Kesan, n=5; Malkara, n=2). Ovitraps were positioned on the ground in green, shady, and easily accessible areas, with a free space of at least 1 meter above, and checked at biweekly intervals (Albieri et al. 2010, Carrieri et al. 2011). Exact altitude and coordinates of sampling stations, and numbers of ovitraps set per site are given in Table 1. Location coordinates and altitudes were estimated using a Magellan® Explorist 5100® GPS receiver.
Ovitraps comprised black plastic cylindrical containers (diameter 11.2 cm, height 11.8 cm, volume 1.1 L). Each contained a single Masonite-like oviposition strip (150×23 mm) and 600 mL of water containing Bacillus thuringiensis israelensis Vectobac® 12AS formulation (stock=1200 International Toxic Units [ITU]/mg) at a dose of 1 mL/L (equivalent to 1279 ITU/mL) to avoid possible larval development according to the protocol of Carrieri et al. (2009). Ovitraps were checked biweekly; oviposition strips were transferred into labeled, airtight plastic bags and transported to the laboratory in a temperature-insulated cooler box without ice. Oviposition strips were individually examined for mosquito eggs using a stereomicroscope in the laboratory.
To facilitate morphological identification, collected eggs collected were reared through to final instar larvae and adults following the published protocol of Medici et al. (2011). Eggs were collected from the oviposition strips using soft brushes, transferred onto filter paper, and submerged into labeled containers filled with dechlorinated water. A total of 4 mg of crushed dry cat food (Friskies®) was provided per larva for the whole larval development, with 10% given on day 0, 45% on day 2, and the rest on day 5. Specimens that died as immatures were stored individually in Eppendorf® tubes containing 96% ethanol at −20°C prior to molecular analysis. Adults were preserved dry at −20°C for later morphological species confirmation.
Morphological identification
Morphological identification of adult male and female mosquitoes was carried out using the available keys of Schaffner et al. (2001) and Becker et al. (2003). Egg characters were compared to those highlighted in Linley (1989).
Molecular identification
DNA was extracted on the QIAgen® BioSprint robotic platform using the BioSprint 96 DNA Tissue Kit (QIAgen®, Crawley, England, UK), following the manufacturer's instructions. The barcode region of the mtDNA cytochrome c oxidase I (COI) gene (658 bp, less primers) was amplified using the universal LCO and HCO barcoding primers (Folmer et al. 1994) using the standard Mosquito Barcoding Initiative (MBI) protocols, expressly listed in Ruiz et al. (2010). PCR products were purified using the Millipore® vacuum manifold system, following the manufacturer's instructions. Bidirectional DNA sequences were generated in the Sequencing Facility of the Natural History Museum, London, using the Big Dye® Terminator Kit (PE Applied BioSystems®, Warrington, England) and run on an ABI 3730 automated sequencer (PE Applied BioSystems®).
Sequences were edited using Sequencher™ version 4.8 (Genes Codes Corporation, Ann Arbor, MI) and compared with published sequences in GenBank and other unpublished sequences in the Mosquito Barcoding Initiative Dataset in the Barcode of Life Database (BOLD,
Voucher DNA extracts are stored in the long-term frozen collection of the Natural History Museum, London (BMNH), where they can be accessed on request. Bidirectional edited COI electropherograms and specimen details of the three Turkish St. albopicta specimens sequenced (including exact localities with georeferences and specimen identifiers) are freely available in the Mosquito Barcoding Initiative project STHAW (Stegomyia of Turkey) in the BOLD database (Ratnasingham and Hebert 2007), and appear in GenBank as barcode red flag data, indicating their high quality and voucher standards under the accession numbers JQ412504–JQ412506. Morphological vouchers, as larvae and adults of both sexes, are held in the laboratory of the senior author (K.O.) and are available for examination on request.
Results
Ovitrap surveillance carried out in 15 sampling stations in the Thrace Region of Turkey from May to October 2011 yielded 4872 mosquito eggs (Table 2). More than 90% of all the eggs collected (n=4828) were those of Culex pipiens sensu lato (s.l.), and the remaining 44 eggs were identified as those of the genus Stegomyia. Although the ovitrap surveillance began in May, 2011, the first Stegomyia eggs were detected in Ipsala (site I6; n=20 eggs) in August, 2011 (week 32) (Table 2, Fig. 2b,c).

Photographs of the two ovitrap locations where St. albopicta eggs were detected in this study: (
DNA barcoding was employed to verify the identity of four small larvae preserved in ethanol from site I6 in Ipsala. DNA fragments corresponding to 658 bp of the mitochondrial COI region were amplified and sequenced from three of these specimens and compared with DNA from specimens of all four species of Stegomyia previously detected in Europe: St. aegypti, St. albopicta, St. cretina, and St. japonica, publicly available in GenBank, or generated through the efforts of the MBI (Fig. 3). The common COI haplotype from these individuals was100% identical with St. albopicta GenBank entry JQ004525 (Fig. 3), thus confirming the presence of St. albopicta in Turkey for the first time.

Bootstrap neighbor-joining tree constructed using 1000 replicates of Kimura two-parameter distance model genetic distance matrices (Kimura 1980) of cytochrome c oxidase sequences (658 bp) belonging to 16 specimens of the four Stegomyia species previously reported in Europe; Stegomyia albopicta (n=7; 3 from Turkey herein, HQ398900-1, JQ004524-5), St. aegypti (n=4, herein), St. cretina (n=3, herein), and St. japonica (n=2; HQ978777-8). The optimal tree with the sum of branch length=0.26529964 is shown. The percentages of replicate trees in which the associated taxa clustered together in the bootstrap test are shown next to the branches (Felsenstein 1985). The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. Evolutionary analyses were conducted in MEGA v.5.0 (Tamura et al. 2011).
The COI sequences of all three specimens of St. albopicta sequenced from ovitrap I6 were identical, indicating that these individuals may well represent siblings from the same mother (i.e., one egg batch) (Table 3). Intraspecific variation was low (<0.1%) in St. albopicta (n=7), St. aegypti (n=4), and St. cretina (n=3), and slightly higher (0.8%) in St. japonica (n=2). Genetically, St. albopicta is most closely related to St. cretina (11.5% sequence difference), than St. aegypti (13.8%) and St. japonica (14.5%) (Table 3, Fig. 3)
Subsequent and closer inspection of fourth instar larvae and the 12 adult specimens reared from the ovitrap at site I6 also confirmed the presence of St. albopicta by morphology and served to provide pristine voucher specimens of this alien taxa. All Stegomyia eggs detected in this study were reared to optimize correct morphological verification.
Eggs of St. albopicta were obtained in two discrete localities in Kesan (ovitrap K5) and Ipsala (ovitrap I6) districts of the Thrace Region of western Turkey, some 30–60 km from the Turkish-Greek border (Fig. 1, Tables 1 and 2). In Ipsala, 20 St. albopicta eggs were collected from an ovitrap set in shrubbery near to the Ipsala border gate (I6: Figs. 1 and 2b,c) in week 32 and a further 6 eggs collected from the same ovitrap in week 38 (Table 2). One other positive St. albopicta collection (n=18 eggs) was from an ovitrap set in the grounds of a restaurant in Kesan (site K5; Figs. 1 and 2a), 30 km east of Ipsala City, in week 36 (Table 2). At sites K5 and I6, St. albopicta eggs were collected in sympatry with egg rafts of Cx. pipiens s.l. That St. albopicta eggs were collected in two localities in the Thrace Region of Turkey, some 30 km distant from each other, and at the same site in Ipsala 6 weeks apart, suggests that St. albopicta is not only present in northwestern Turkey, but has in fact established low-level populations in this area.
In a concurrent survey, three adult females Stegomyia were detected a gravid trap in vegetation in Ipsala City, on July 12, 2011. Although these specimens were tentatively identified as St. cretina in the field, we are now more inclined to believe that these specimens were also St. albopicta. No other St. cretina specimens were detected in this study.
In this study, eggs of Cx. pipiens s.l. were collected from almost all sampling stations (n=4828 in 67 egg rafts) (Table 2). During first week of trapping (week 23), no egg rafts were collected, and in the subsequent 2 weeks, collections were occasional. However from weeks 26 to 38, collections became consistent and more intense. Among the sampling stations, K5 in Kesan and I8 and I6 in Ipsala yielded the most Cx. pipiens s.l. eggs, with totals of 968, 743, and 638 eggs, respectively. The population size of Cx. pipiens s.l. showed unimodal increase. During the study period, the population of these mosquito species was found to be lowest in June (n=141). Population size increased during July (n=1379) and August (n=1784), with the highest peak in August (week 34), and decreased during September (n=1430) and October (n=227).
Discussion
Herein, the presence of St. albopicta was confirmed for the first time in western Turkey in August, 2011, through retrospective correlation of 658-bp mitochondrial DNA COI gene sequences (“DNA barcodes”) with those of morphologically verified specimens sequenced by the MBI. DNA barcoding was employed to facilitate identification from early larval instars (1–3), which cannot reliably be identified using taxonomy alone. Given that potentially invasive taxa are rarely included in most commonly used regional keys, we advocate the molecular confirmation of newly encountered invasive species to conclusively confirm the identity of alien taxa (Scholte et al. 2007 and herein). Once the identity of an alien taxa has been confirmed, morphological identifications can be employed for routine screening.
The prediction that “the climatic suitability of Turkey would dictate that it is only a matter of time before the species is introduced and established” (Unlu and Farajollahi 2012) and an earlier ECDC report (2009) directly led to the implementation of this active surveillance study in western Turkey and clearly shows the value of predictive risk modeling of potentially invasive vector species in previously identified localities (Mitchell 1995b, Knudsen et al. 1996, Benedict et al. 2007, ECDC 2009, Fischer et al. 2011, Caminade et al 2012, Unlu and Farajollahi 2012). Along with Turkey, Bulgaria and southern Russia were highlighted to be at high risk of invasion, and the presence of St. albopicta in these zones was reported at a scientific meeting in 2011 (see Introduction) and in early 2012 (Anonymous 2012), respectively. Given that these predictions were valid in Bulgaria, other countries highlighted in these previous studies (Cyprus, Macedonia, Portugal, and southern Russia) should take these warnings seriously and undertake active surveillance programs (ECDC 2009, Unlu and Farajollahi 2012). This study supports the effectiveness of biweekly ovitraps for the detection of St. albopicta, as employed in Italy (Albieri et al. 2010, Carrieri et al. 2011), and we herein support this methodology as a simple cheap and effective method for active surveillance of this invasive species. That Cx. pipiens s.l. eggs were collected in the ovitraps indicate that this method can also be used to effectively monitor Cx. pipiens s.l. populations, perhaps in a combined virus vector surveillance program in future.
That St. albopicta is reported in western Turkey herein for the first time is of significance to public health authorities in Turkey. The ability of St. albopicta to use both natural and artificial containers for larval habitats facilitates its widespread occupation of urban and periurban environments (Hawley 1988), ensuring a close connection between the species and the human population, increasing the risk of vector-borne diseases in these areas (Takken et al. 2007). The spatial and temporal confirmation of the species in two different locations of the Thrace Region, 30–60 km from the Greek–Turkish border in this study, indicates that not only is the species present in the country, but also that indeed the species may well be established in this region of Turkey, albeit at low population density at the current time.
St. aegypti was reported in the Aegean port city of Izmir and nearby Odemis in Turkey in 1961, but this was believed to be an accidental importation and the species did not establish in this region (Curtin 1967). St. cretina, native to Greece, is the only species of the genus native to Europe, yet invasions of St. albopicta, St. aegypti, and St. japonica have all been reported (Curtin 1967, Adhami and Murati 1987, Benedict et al. 2007, Scholte et al. 2007, Gatt et al. 2009, ECDC 2009, Schaffner et al. 2009, Anonymous 2012), and the latter three predicted in Turkey (ECDC 2009, Unlu and Farajollahi 2012). Herein we show that DNA barcodes readily separate St. albopicta, St. aegypti, St. cretina, and St. japonica and this technique should prove useful in determining the true identity of invasive Stegomyia species detected across Europe in future.
Geographic information systems (GIS) models recently presented by Caglar and Karacaoglu (2011) showed the possible expansion pattern of the species in Turkey, indicating that most regions of the country were suitable for occupation by St. albopicta; thus, efforts must be undertaken to prevent the establishment of the species. Early detection of St. albopicta through active surveillance coupled with targeted suppression measures to prevent establishment is promoted as the most cost-effective method of St. albopicta control (Mack et al. 2000, Derraik 2006, ECDC 2009). Conventional larval control strategies (e.g., insecticide spraying, larval site management) have proven highly ineffective in controlling already established, high-density St. albopicta populations due to the dispersal and availability of larval sites (Mack et al. 2000, Derraik 2006). Due to typical discontinuous distribution and low active dispersal potential and ease of mass-rearing (Bellini et al. 2007), one of the most promising control measures to date is the sterile insect technique (SIT), which requires sustained effort and funding.
Governmental-level policy to avoid the invasion of etiological agents and the insects that carry them is generally underdeveloped in Europe, reflecting the low level of threat to which we have previously been exposed. Island countries, including Australia and New Zealand, are extremely tight on border controls to avoid accidental importation of insect disease vectors and agricultural pests, and these countries maintain well-established early warning systems including active surveillance. Implementation of a program to prevent the establishment of St. albopicta and associated viruses/parasites transmitted by the species in Turkey was suggested by Ergunay et al. (2011).
Given the detection of St. albopicta in Turkey's western Thrace Region herein, we advocate that active surveillance must now be implemented not only in northwestern Turkey, but also in other regions of the country that are at risk. St. albopicta has been reported in Latakia, Syria, which is only 60 km south from the Yayladagı border gate of Hatay, Turkey (Haddad et al. 2007). The Turkish–Syrian border has 12 border gates along the frontier, with the Yayladagı border gate facilitating one of the major immigration and trade routes through the Hatay province. Turkey is also at risk from passive introduction of mosquitoes in vehicles coming from neighboring countries with established populations of St. albopicta (Bulgaria, Greece and Syria), onboard boats coming from Greece and Italy, and as desiccated eggs in used tire importations.
Although border zones are key to the introduction of invasive species, the role of internal transportation (of both the mosquitoes and the diseases they transmit) via human movement should not be overlooked. Internal movements of large numbers of agricultural workers and laborers between South Eastern Anatolia and the Cukurova plain and areas in the Mediterranean, Aegean, and Thracean provinces could result in these persons returning to the central plateau and eastern highlands, carrying the vector-borne pathogens and some of the vectors with as well (Alten et al. 2007, Takken et al. 2007).
Our confirmation of St. albopicta in Turkey coupled with the recent detection of dengue seroactivity in humans in Ankara (Ergunay et al. 2010) compounds the need for active detection and elimination practices of this species in Turkey. Further investigations will be conducted during the 2012 season to determine the extent of this incursion in Turkey, survey additional areas and actively initiate control measures.
Conclusions
This study documents the presence of St. albopicta in Turkey for the first time in August, 2011, and supports the strategy of active surveillance programs in areas identified as high risk through geographical, climatic, and ecological niche modeling. DNA barcoding has proven an accurate diagnostic method of confirming the identity of early instar St. albopicta and differentiating it from other morphologically similar invasive species in Europe (St. aegypti, St. cretina, and St. japonica), thus we advocate this method for the rapid confirmation of alien taxa in future active ovitrap surveillance surveys.
That St. albopicta has been detected in low density in western Turkey implies that targeted vector control strategies can still be employed in the region to avoid the establishment of high-density populations of this effective arboviral vector. Given the serious diseases St. albopicta is known to transmit to humans, prevention of the establishment of this species should be regarded as a high priority for public health authorities in Turkey.
Footnotes
Acknowledgments
Funding for this study was provided by the Scientific Research Projects Coordination Unit of Istanbul University (Project Number 11459) and Hacettepe University Scientific Research Unit, Ankara, Turkey. DNA sequencing for this study was carried out as part of the activities of the Mosquito Barcoding Initiative (Project Leaders: Y.M.L. and Richard C. Wilkerson, Walter Reed Biosystematics Unit [WRBU], Smithsonian Institution, USA). We are grateful to MBI collaborators, Major Joshua Bast (USAMRU-Kenya) and Dr. G. Koliopoulos (Benaki Institute, Athens, Greece), for providing specimens used as outgroups in this study, and to Dr. Desmond Foley (WRBU) and Ssgt. Luke Mitchell (British Army) for assistance with the map figure. This manuscript was prepared while Y.M.L. held a National Research Council (NRC) Research Associateship Award at the Walter Reed Army Institute of Research. The material to be published reflects the views of the authors and should not be construed to represent those of the US Department of the Army or the US Department of Defense.
Author Disclosure Statement
No competing financial interests exist for any of the authors involved in this manuscript.