
Abstract
Rodents and soricomorphs are animal hosts of fleas and associated zoonotic microbial pathogens. A total of 4,889 small mammals were collected from Gyeonggi and Gangwon Provinces, Republic of Korea, from 2008 through 2010, including: Apodemus agrarius (4,122, 84.3%), followed by Crocidura lasiura (282, 5.8%), Microtus fortis (257, 5.3%), Myodes regulus (77, 1.6%), Micromys minutus (71, 1.5%), Mus musculus (63, 1.3%), and 4 other species (17, 0.3%). A total of 1,099 fleas belonging to 10 species and 7 genera were collected. Ctenophthalmus congeneroides (724, 65.9%) was the most commonly collected flea, followed by Stenoponia sidimi (301, 27.4%), Neopsylla bidentatiformis (29, 2.6%), and Rhadinopsylla insolita (25, 2.3%). The remaining species accounted for only 1.8% (20, range 1–6) of all fleas collected. The 2 dominant flea species, C. congeneroides and S. sidimi, showed an inverse seasonal pattern, with higher populations of C. congeneroides from January–September, whereas S. sidimi was more frequently collected during October–December. The overall flea infestation rates (FIR) and flea indices (FI) were 14.1% and 0.22, respectively, and were highest during April–June (19.7% and 0.30, respectively). A total of 735 of the 1,099 fleas were assayed for the detection of Bartonella spp. by PCR using Bartonella-specific primers, of which 515 were positive for Bartonella, with an overall maximum likelihood estimate (MLE) of 700.7/1,000. The highest MLE values were observed during April–June (899.2) and July–September (936.2) trapping periods and, although lower, were similar for January–March (566.7) and October–December (574.1). C. congeneroides demonstrated high MLEs for all seasons (range 752.5–934.8), while S. sidimi was positive for Bartonella only during January–March (MLE=342.1) and October–December (MLE=497.2) collection periods. Continued long-term surveillance of small mammals and associated ectoparasites is needed to improve our understanding of the prevalence of Bartonella spp. in fleas and the role of fleas in the zoonotic maintenance and transmission of Bartonella to humans.
Introduction
In the Republic of Korea (ROK), rodents and soricomorphs are animal hosts to a number of zoonotic pathogens (i.e., Rickettsia, Ehrlichia, Anaplasma, Bartonella, and Borrelia spp.), whereas fleas, ticks, and mites serve as vectors (Park et al. 1992, Kee et al. 1994, Chae et al. 2003, Lee et al. 2003, Kim et al. 2005). A survey of fleas collected from rats was first reported in Korea by Kobayashi (1931). In subsequent surveys, many flea species were observed and associated with natural hosts (Nagahana 1937, 1938, 1954, Tipton et al. 1972, Hong 1994). Recently, a preliminary investigation of zoonotic bacterial pathogens showed that the prevalence of Bartonella spp. from ticks, mites, and tissues of small mammals was relatively high, compared to other bacterial pathogens (Kim et al. 2005). In addition, Rickettsia typhi and R. felis were detected by PCR in fleas collected from rodents and soricomorphs (Kim et al. 2010, Ko et al. 2011).
Members of the genus Bartonella are Gram-negative obligate bacteria of mammalian red blood cells. Some species of Bartonella are the etiological agents of human and animal diseases that vary from mild-to-severe febrile illnesses, including endocarditis, neuroretinitis, and bacillary angiomatosis (Tappero et al. 1993, Kerkhoff et al. 1999, Tsukahara et al. 2000, Fenollar et al. 2005, Morway et al. 2008). There have been many studies related to the identification of Bartonella spp. in rodents worldwide. On the basis of these and other studies, it was shown that flea control resulted in the reduction in the prevalence of Bartonella in rodents, supporting the importance of their role in the maintenance and transmission of Bartonella bacteria among host populations and the potential for incidental transmission to domestic animals and humans (Jardine et al. 2006, Morway et al. 2008). In support of this, Bartonella species that were associated with rodent hosts also were observed to be associated with illnesses in humans (Welch et al. 1999, Comer et al. 2001, Smith et al. 2002, Kosoy et al. 2003, Iralu et al. 2006, Morway et al. 2008). Therefore, vector- and rodent-borne disease surveillance is critical to identify associated health threats and risks to reduce the potential for transmission of these zoonotic pathogens to humans.
Ultimately, comprehensive infectious disease surveillance programs that identify the diversity of hosts and their associated ectoparasites and the potential for human exposure to zoonotic vector-borne disease agents will provide a better understanding of disease maintenance cycles in nature and for predicting the emergence of infectious diseases and transmission to man (Nieto et al. 2007). Annual environmental factors, especially reforestation and rainfall patterns, and warming annual climatic trends over the past few decades, affect host and associated ectoparasite populations (Chilton et al. 2000, Chae et al. 2008). Therefore, multiple-year surveillance of hosts, ectoparasites, and associated pathogens is essential to the understanding of zoonotic pathogen maintenance cycles and predicting the emergence of infectious diseases of veterinary and medical importance.
In this study, fleas were collected from small mammals and assayed by PCR for the detection of Bartonella to determine host associations, seasonal and annual prevalence of fleas, and the maximum likelihood estimations (MLE) of Bartonella in fleas collected from small mammals from 2008 through 2010.
Materials and Methods
Samples and sampling sites
Fleas were collected from small mammals (rodents and soricomorphs) captured at US military installations and US- and ROK-operated training sites in Gyeonggi (Paju, Pocheon, Osan, Suwon, and Pyeongtaek) and Gangwon (Cheolwon) Provinces from March, 2008, through December, 2010 (Fig. 1). Small mammal trapping was conducted at rice paddies, military cantonment and fighting position sites, training area perimeters, intermittent streams, ponds, and areas that were infrequently traveled by vehicles or used for troop patrols and vehicular maneuver activities (Kim et al. 2011). Small mammal trapping was conducted as part of the US military hantavirus surveillance program, which consisted of surveying rodent-infested habitats within military sites. The primary vegetation consisted of tall grasses with various proportions of herbaceous vegetation and shrubs, and small groves of young deciduous trees with grasses and shrubs as understory (Kim et al. 2011). Surveys were conducted quarterly from January–March, April–June, July–September, and October–December. Collapsible live-capture Sherman® traps (7.7×9×23 cm; H.B. Sherman, USA) baited with crackers and peanut butter, were set out during the daytime and checked the following morning. Nonabsorbent cotton balls were placed in the traps during spring and winter months to maintain small mammal temperatures and therefore reduce their mortality due to low nighttime temperatures. Traps positive for small mammals were numbered sequentially, placed in secure shipping containers, and transported to Korea University, Seoul. Small mammals were given a unique number, anesthetized, identified to species by morphological methods, sexed, and euthanized by cardiac puncture in accordance with Korea University animal use protocol (O'Guinn et al. 2010). For each small mammal, fleas were removed and placed individually in cryovials containing 100% ethyl alcohol. Each cryovial was labeled sequentially by host, and then transported to the 5th Medical Detachment, 168th Multifunctional Battalion, 65th Medical Brigade, Yongsan Army Garrison, Seoul, where they were identified to species and sex under a stereomicroscope using conventional taxonomic keys (Hopkins and Rothschild 1953, 1956, Hong 1994).
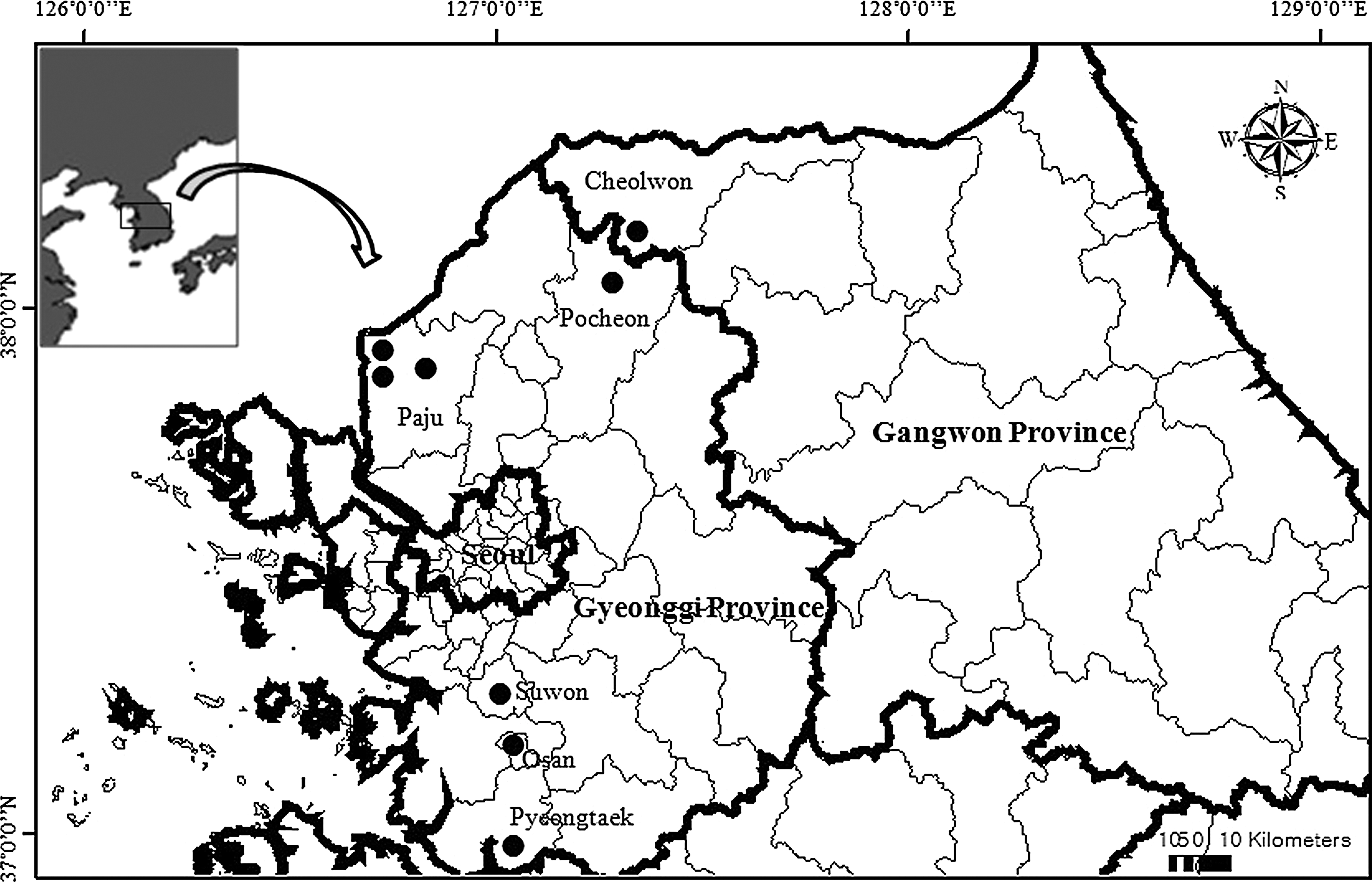
Small mammal collection sites (dots) in Gyeonggi and Gangwon Provinces, Republic of Korea from 2008 through 2010. General site locations were: Warrior Base, Twin Bridge Training Areas and Dragon Head (Paju City), Nightmare Range (Pocheon County), Osan Air Base (Osan City), Suwon Air Base (Suwon City), and Camp Humphreys (Pyeongtaek City) in Gyeonggi Province; Chipori Range (Cheolwon County) in Gangwon Province.
DNA extraction and PCR
Total genomic DNA was extracted from 735 individual fleas (n=250, 235, and 250 for 2008, 2009, and 2010, respectively) for the detection of Bartonella-specific DNA. Each flea was homogenized using a sterile Beadbeater TissueLyser II (Qiagen, Germany) with 180 μL of lysis buffer, 20 μL of proteinase K, and 5-mm stainless steel beads at 30 frequencies/s for 5 min, incubated overnight at 56°C, and then centrifuged at 20,000×g for 15 min at room temperature. After centrifugation, the supernatant was used for DNA extraction using DNeasy Tissue Kits (Qiagen, Germany). Two sequential PCRs for the detection of the RNA polymerase β-subunit (rpoB) gene for Bartonella were conducted using 2 sets of primers (Table 1). The PCRs were carried out in a 25-μL reaction volume containing 10–100 ng for the 1st PCR and 1 μL of the 1st PCR product for the 2nd PCR of DNA template, 10×PCR buffer (Takara, Korea), 2.5 mM of each deoxyribonucleotide triphosphate (dNTP), 10 pmol of primers, and 1 unit DNA Taq polymerase (Takara, Korea). PCR was performed in a PTC-200 thermal cycler with the following conditions: 1st PCR, initial denaturation for 5 min at 94°C, followed by 30 cycles (94°C for 45 s, 57°C for 45 s, and 72°C for 45 s) with a final extension for 5 min at 72°C; 2nd PCR, initial denaturation for 5 min at 94°C, followed by 25 cycles (94°C for 30 s, 60°C for 30 s, and 72°C for 30 s) with a final extension for 5 min at 72°C. The PCR products were resolved by electrophoresis on a 1.5% agarose gel and detected using ethidium bromide.
Annealing temperature in PCR reactions.
Cloning, sequencing, and phylogenetic analysis
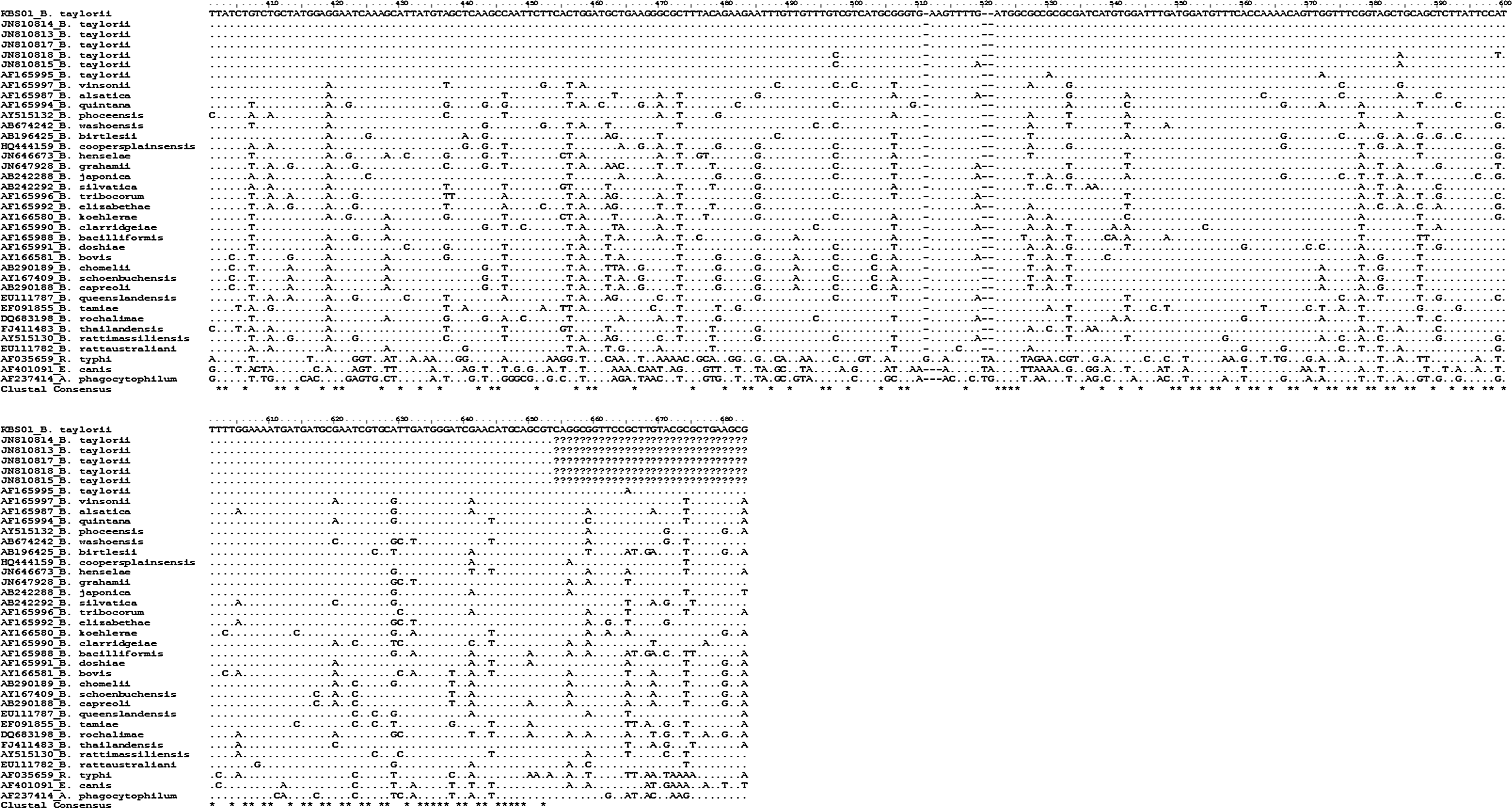
PCR products were purified with QIAquick Gel Extraction kits (Qiagen, Germany), and then cloned with pGEM®-T Easy Vectors (Promega Corp., USA) according to the manufacturer's protocols. Plasmid DNA for sequencing was again purified using the Wizard® Plus SV Minipreps DNA Purification System (Promega Corp., USA). Purified recombinant plasmid DNA was sequenced using T7 and SP6 promoter primers with an automatic sequencer (ABI 3730xl capillary DNA sequencer, USA). All sequences were manually edited at least 5 times on AlignIR program version 2.1 (LI-COR Inc., USA). Analyses of the sequences were comparatively completed using a BLAST search for Bartonella rpoB sequences in GenBank. A multiple sequence alignment was conducted using BioEdit version 7.0.9.0 (Fig. 2; Hall 1999). The phylogenetic relationships of the rpoB sequences were constructed using the neighbor-joining method (Saitou and Nei 1987) under the Kimura 2-parameter mode (Kimura 1980) (Fig. 3). As reference sequences, corresponding rpoB sequences were chosen from GenBank database (Figs. 2 and 3). Also, corresponding rpoB sequences of Rickettsia typhi, Ehrlichia canis, and Anaplasma phagocytophilum were selected to root the phylogenetic tree as outgroups. Confidence in estimated relationship was determined using the bootstrap approach (Felsenstein 1985) obtained through 1,000 replicates incorporating the same model as above. Bootstrap analysis and phylogeny reconstruction were conducted using MEGA version 4.0 (Tamura et al. 2007).

Summary of partial rpoB gene sequences of Bartonella spp., with corresponding rpoB sequences of Rickettsia typhi, Ehrlichia canis, and Anaplasma phagocytophilum. ‘·’ and ‘?’ mean the identical and unknown nucleotide sequences, respectively.
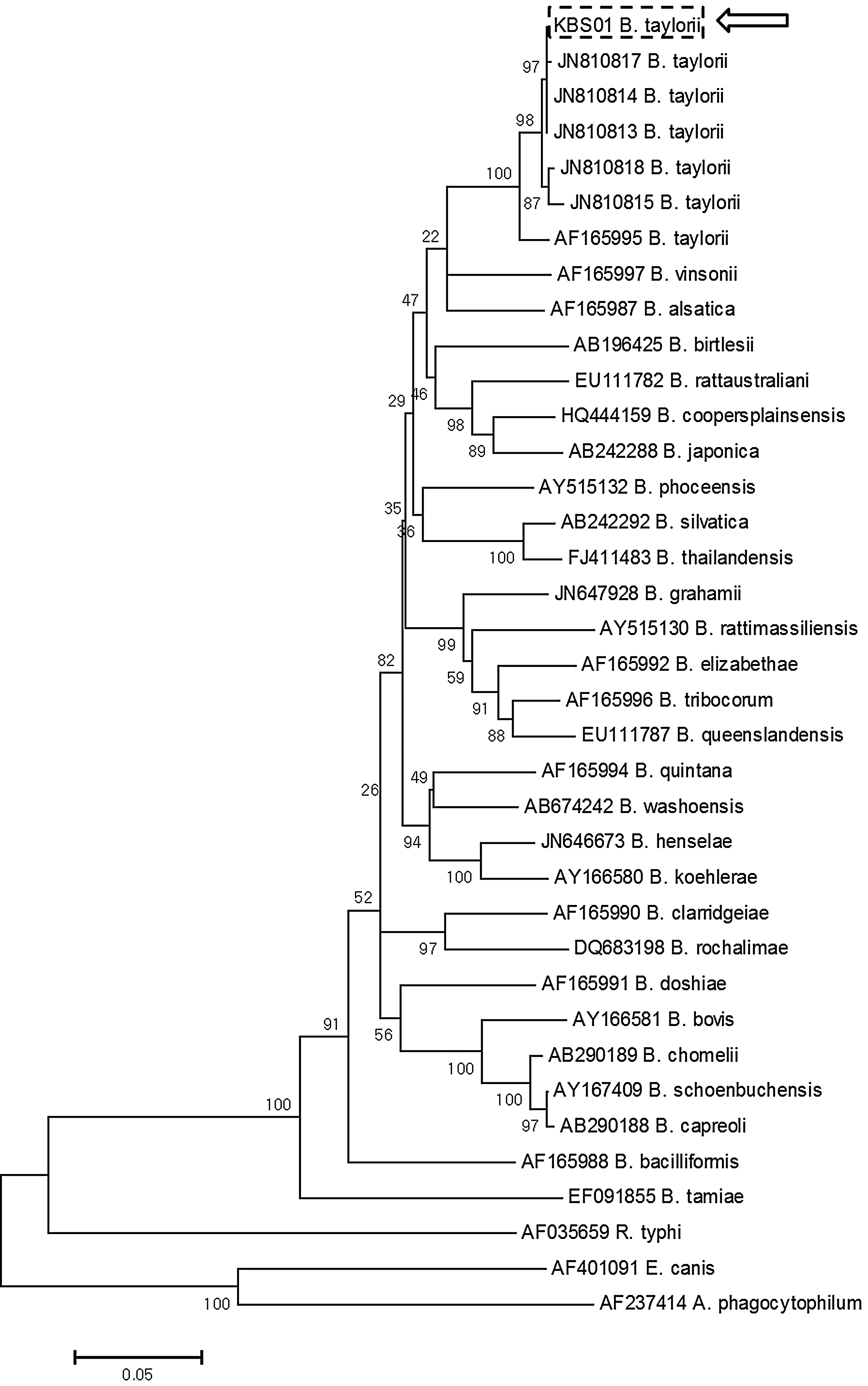
The phylogenetic tree of partial rpoB gene sequences of Bartonella taylorii, and the other Bartonella spp. As outgroups, corresponding rpoB sequences of Rickettsia typhi, Ehrlichia canis, and Anaplasma phagocytophilum were used. The tree was rooted using 3 different sequences. The arrow indicates a genotype (KBS01 B. taylorii strain) detected in this study.
Data analyses of prevalence
Flea infestation rates (FIR) and flea indices (FI) were calculated (Kim et al. 2010, Ko et al. 2011) as follows: FIR=(Number of mammals, by species, with fleas/Total number of mammals, by species)×100; FI=Number of fleas collected from mammals, by species/Total number of mammals, by species. The MLE infection rates were calculated using the PooledInfRate Software version 4.0 (Biggerstaff 2006). In the traditional analysis of prevalence data using the MFIR, an assumption is required that when a pool is positive, then only 1 individual in that pool is positive. However, MLE methods do not require this assumption. Although we used 1 flea for each pool, the MLE method was applied in this study. For more detailed information, refer to Biggerstaff (2006).
Results
A total of 1,099 fleas were collected from 4,889 small mammals belonging to 10 species and 4 genera (Table 2). Apodemus agrarius (4,122, 84.3%) was the most frequently captured small mammals, followed by Crocidura lasiura (282, 5.8%), Microtus fortis (257, 5.3%), Myodes regulus (77, 1.6%), Microtus minutus (71, 1.5%), and Mus musculus (63, 1.3%), whereas the remaining 4 species accounted for only 0.3% (17) of all small mammals collected. Overall, the FIR was 14.1%, ranging from 3.2% (M. musculus) to 66.7% (Tamias sibiricus) (Table 2). The overall FI for all mammals was 0.22 and highest from T. sibiricus (1.33), followed by A. peninsulae (0.50) and Rattus norvegicus (0.33) (Table 2). The overall mean number of fleas collected from the small mammals was 1.6 (range 1.0–2.0) (Table 2).
Mean number of fleas per infested mammal for each species (no. fleas collected/no. mammals with fleas).
Overall, the number of fleas was similarly collected during October–December (355, 32.3%), April–June (335, 30.5%), and January–March (312, 28.4%), whereas lower numbers were collected during July–September (97, 8.8%) (Table 3). For sites collected during all seasons and years (Paju, Pocheon, and Pyeongtaek), the FIRs and FIs were highest for Paju (21.1%, 0.35), whereas they were similar for Pocheon (12.9%, 0.19) and Pyeongtaek (12.5%, 0.21). Both FIRs and FIs for all fleas were similar and highest during April–June (19.7%, 0.30), October–December (17.4%, 0.28), and January–March (15.4%, 0.25) but much lower during July–September (4.4%, 0.08) (Fig. 4).
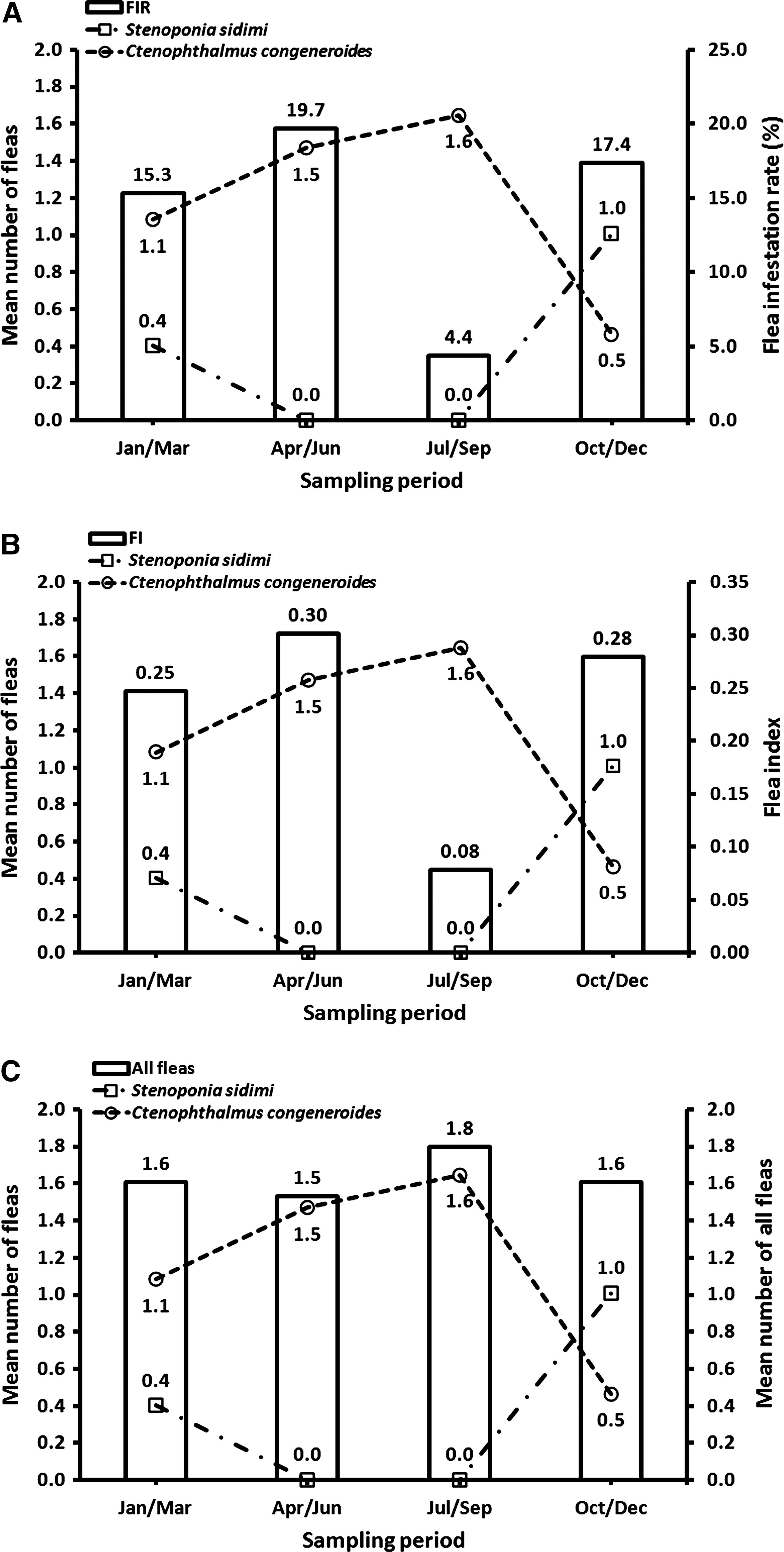
Flea infestation rates (FIR) (
Mean number of fleas per infested mammal for each species (no. fleas collected/no. mammals with fleas).
The number of small mammals captured.
Ctenophthalmus congeneroides (724, 65.9%) was the most commonly collected flea from small mammals, followed by Stenoponia sidimi (301, 27.4%), Neopsylla bidentatiformis (29, 2.6%) and Rhadinopsylla insolita (25, 2.3%), whereas the remaining 6 species accounted for only 1.8% (20, range 1–6) of all fleas collected (Table 4). Except for Doratopsylla coreana and Monopsyllus anisus, all species were recorded from A. agrarius (Table 4). The 2 dominant flea species, C. congeneroides and S. sidimi, showed an inverse seasonal pattern, with higher populations observed for C. congeneroides during January–September, whereas S. sidimi was more frequently collected during October–December collection periods (Fig. 4).
Using Bartonella genus-specific PCR assays, Bartonella was detected in 515/735 (70.1%) of the fleas assayed. For the 2 most frequently collected species of fleas, Bartonella was detected in 84.7% (392/463) and 44.9% (115/256) of C. congeneroides and S. sidimi, respectively (Table 5). Overall, the highest MLE rates were observed during July–September (936.2), followed by April–June (899.2), October–December (574.1), and January–March (566.7). The MLEs of C. congeneroides were higher than that of S. sidimi for each of the survey periods. Overall, A. agrarius accounted for the greatest number of fleas collected (962/1099, 87.5%) (Table 4) and largest number of Bartonella-positive fleas (476/515, 92.4%).
One flea per pool (totally 735 fleas or pools).
The number of fleas tested for detection of Bartonella spp.
Not applicable (N/A) in the MLE method.
Overall, annual FIRs for 2008–2010 were similar (range 12.6–20.6%), with FIs ranging from 0.19 to 0.35 (Table 6). The percentage of variation in the Bartonella-positive fleas (58.0–80.4%) was likely affected by differences in collection sites for each of the years and numbers of small mammals collected at each of the sites. In addition, it is difficult to extrapolate our findings to the rest of the ROK, due to geographic limitations of the present samplings. Latitudinal and longitudinal survey site surveillance in the ROK is needed to provide more accurate information of seasonal and annual differences in FIRs, FIs, and MLEs as a result of climatic differences (i.e., rainfall) for Bartonella-positive fleas.
One genotype was detected out of 3 different samples (2 S. sidimi and 1 C. congeneroides collected from A. agrarius in Gyeonggi Province; specimen voucher no. F09-007, 008, and 020) in the study. The sequence was deposited in GenBank (accession no. JX984664; strain KBS01). The 678-bp rpoB gene nucleotide sequences of the 3 samples were identical to B. taylorii (JN810813-4 with 95% query coverage). A consensus phylogenetic tree was reconstructed with reference and outgroup sequences (Fig. 3). To date, 5 species, B. taylorii (S. Ko, unpublished data), B. grahamii (S. Ko, unpublished data), B. henselae (Kim et al. 2009), B. clarridgeiae (Kim et al. 2009), and B. elizabethae (Kim et al. 2005), have been detected in the ROK.
Discussion
At the end of the Korean War (1950–1953), the Korean landscape was largely devoid of trees due to military activities and poverty, which restricted the development of small (i.e., rodents, soricomorphs, and associated predators), medium (i.e., raccoon dogs and leopard cats), and large (i.e., water deer and wild boar) mammal populations that use forests for refuge. During the 1960s, a national tree planting policy was instituted by the ROK government, with nonagricultural lands (hills and mountains) now encompassing large expanses of groves of planted trees, i.e., chestnuts, oaks, pines, larch, and volunteer trees (Lee and Lee, 2002). As Korea consists of 70% mountainous terrain, this greatly increased the potential for harborage of wild mammals, i.e., water deer, wild boar, raccoon dogs, leopard cats, weasels, rodents, soricomorphs, and feral cats and dogs, and the associated ectoparasites, i.e., fleas, ticks, and mites, that they harbor. Recently, R. typhi (Typhus group) and R. felis (Spotted fever group Rickettsia) were detected in fleas collected from small mammals in the ROK (Kim et al. 2010, Ko et al. 2011). However, identification and distribution of flea-borne pathogens, including Bartonella, has not been well documented in the ROK.
During recent years, a number of Bartonella species, which are etiological agents of many human and animal diseases, have been isolated and characterized, including a few model organisms for studying the evolution and ecology of vector-borne diseases (Jacomo et al. 2002). For examples, sandfly-borne B. bacilliformis (oroya fever) in a Peruvian Andean village, South America (Boulouis et al. 2005), louse-borne B. quintana (trench fever) in many countries of the world (Fournier et al. 2002), flea-borne B. henselae (cat scratch disease and bacillary angiomatosis) (Karem 2000), and other Bartonellae in humans. However, few studies have been conducted on Bartonella species in the ROK, compared with the numerous studies through the world. Although some of reports on the molecular detection of Bartonella from fleas of pets (Han et al. 2006), ticks, mites, and small mammals (Kim et al. 2005, Kim et al. 2009), this is the first report on the prevalence of flea-borne Bartonella from small mammals in the ROK.
A. agrarius was the most frequently captured mammal among 10 different small mammal species, and had the most diverse spectrum of fleas (8 of 10 species of fleas, including C. congeneroides and S. sidimi), as previously reported by others (Kim et al. 2010, Ko et al. 2011). These data implicate A. agrarius and associated fleas as hosts/vectors for the maintenance and transmission of Bartonella in the ROK. In general, A. agrarius is found throughout the ROK in a wide range of rural habitats and in tall grass habitats associated with parks and other areas in urban environments (Kim et al. 2011) and is the primary host for Hantaan virus in the ROK (Song et al. 2009, Kim et al. 2011). The habitats in rural farmlands and local and national parks increase the likelihood of human–flea contact, which increases the potential for transmission of flea-borne pathogens to human populations.
The seasonal distribution of FIRs was similar to those observed from previous studies (Walton and Hong 1978, Kim et al. 2010, Ko et al. 2011), except for June (Kim et al. 2010) and August to September (Ko et al. 2011). FIs showed similar seasonal patterns to FIRs, but the mean number of fleas showed an inverse pattern to FIRs. The two most commonly collected flea species, C. congeneroides and S. sidimi, demonstrated distinct seasonal distributions and prevalence of Bartonella. Kim et al. (2010) suggested that this might be due to host and/or flea seasonal reproductive differences. However, there are no clear studies to answer this question. In addition, positive fleas found in this study were observed mainly from fleas of A. agrarius, whereas those of the other species accounted for only minor proportion as a result of relatively low catches for those species.
Occasional flooding, due to seasonal monsoon rains and typhoons, may increase the movement and dispersal of small mammals and associated ectoparasites and zoonotic pathogens that they harbor (Kim et al. 2011). During some years, the rainy season extends from August through September resulting in periodic and occasional flooding of low-lying areas, while during other years the rainy period is much shorter. Also, global climatic change has been reported in the ROK with unknown effects on vegetation, wildlife, domestic animals, and humans. Vectors (i.e., mosquitoes, fleas, lice, ticks, and mites) that harbor infectious agents are sensitive to climatic changes (i.e., increasing temperature, relative humidity, sunshine hour, and precipitation) and may increase/decrease the potential transmission of zoonotic pathogens to domestic animals and humans (Chae et al. 2008). In addition, the accidental or intentional contact with vectors are increasing and pose serious public health risks, particularly as a result of public recreation activities, agricultural and urban expansion, outdoor construction, military training/operations, or possibly, in the event of a natural disaster (Richards et al. 1997, Eisen et al. 2007, Kim et al. 2010, Ko et al. 2011). The present multiple-year prevalence study provides important information on the epidemiology of flea-related Bartonella pathogens in the ROK. However, long-term monitoring of at least 5–10 years and more detailed analyses of relationships between environmental factors, prevalence, distribution of zoonotic flea-borne pathogens, and anthropogenic disturbance are essential for understanding disease maintenance cycles and predicting the emergence of vector-borne diseases affecting veterinary and medical health under changing climatic conditions.
Footnotes
Acknowledgments
We thank the commanders and personnel of the 5th and 38th Medical Detachments, 168th Multifunctional Medical Battalion, for their support. This study was supported by the Armed Forces Health Surveillance Center, Global Emerging Infections Surveillance and Response System, Silver Spring, Maryland, the National Center for Medical Intelligence, Fort Detrick, Maryland, Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2012-0005524), and the National Research Foundation of Korea Grant funded by the Korean Government [Ministry of Education, Science and Technology (NRF-2011-550-20110038)]. The opinions expressed herein are those of the authors and are not to be construed as official or reflecting the views of the US Department of the Army, Department of Defense, or the US Government.
Author Disclosure Statement
No competing financial interests exist.