
Abstract
The detection of Trypanosoma cruzi infection in mammals is crucial for understanding the eco-epidemiological role of the different species involved in parasite transmission cycles. Xenodiagnosis (XD) and hemoculture (HC) are routinely used to detect T. cruzi in wild mammals. Serological methods are much more limited because they require the use of specific antibodies to immunoglobulins of each mammalian species susceptible to T. cruzi. In this study we detected T. cruzi infection by trans-sialidase (TS) inhibition assay (TIA). TIA is based on the antibody neutralization of a recombinant TS that avoids the use of anti-immunoglobulins. TS activity is not detected in the co-endemic protozoan parasites Leishmania spp and T. rangeli. In the current study, serum samples from 158 individuals of nine wild mammalian species, previously tested by XD, were evaluated by TIA. They were collected from two endemic areas in northern Argentina. The overall TIA versus XD co-reactivity was 98.7% (156/158). All 18 samples from XD-positive mammals were TIA-positive (co-positivity, 100%) and co-negativity was 98.5% (138/140). Two XD-negative samples from a marsupial (Didelphis albiventris) and an edentate (Dasypus novemcinctus) were detected by TIA. TIA could be used as a novel tool for serological detection of Trypanosoma cruzi in a wide variety of sylvatic reservoir hosts.
Introduction
The trans-sialidase (TS) is an enzyme expressed by T. cruzi that is not detected in other co-endemic parasites such as Leishmania spp., Trypanosoma rangeli, or Plasmodium spp. (Clough et al. 1996, Frasch 2000). The detection of TS-neutralizing antibodies allowed the development of the TS inhibition assay (TIA), for the diagnosis of T. cruzi chronic infections in humans and naturally infected dogs and cats (Leguizamón et al. 1994, Leguizamón et al. 1998, Buchovsky et al. 2001, Blejer et al. 2008, Sartor et al. 2011). TIA detects TS-neutralizing antibodies by measuring the remnant TS activity after the interaction of serum samples with recombinant TS, thus avoiding the use of anti-immunoglobulins.
In this study we evaluated the performance of TIA in a wide diversity of naturally infected wild mammalian hosts to contribute to the development of a serological tool for T. cruzi detection in the sylvatic transmission cycle.
Methods
Samples
Serum samples from 66 mustelids, 52 marsupials, and 40 edentates of various species (listed in Table 1) previously diagnosed by XD were tested by TIA. Samples were collected by venipuncture from antebrachial, cephalic, saphenous, or jugular veins in field surveys conducted in Amamá (Santiago de Estero Province) between 2003 and 2007 and in Pampa de Indio (Chaco Province) in 2008, both located in northern Argentina. Mammals were captured with a significant effort totaling 7251 National traps-nights and 3467 Sherman traps-nights in Santiago del Estero Province, and 1599 and 440 traps-nights, respectively, in Chaco Province. Serum samples collected were stored at −20°C. We selected a convenience sample from banked samples, including those from all individuals that were positive by XD (5 marsupials, 2 mustelids, and 11 edentates) and a representative number of XD-negative samples for each group of mammals (47 marsupials, 64 mustelids, and 29 edentates) (Ceballos et al. 2006, Alvarado-Otegui et al. 2012).
Xenodiagnosis
Parasitological examination was done following the procedure described elsewhere (Ceballos et al. 2006). Briefly, noninfected, laboratory-reared fourth- or fifth-instar nymphs of Triatoma infestans were exposed to the same individual during 25 min. The number of bugs fed on each mammalian species was 10 for edentates and 20 for marsupials and mustelids. Bug feces were microscopically examined for T. cruzi infection at 30 and 60 days after feeding.
Trans-sialidase inhibition assay
TIA was performed by incubating 15 μL of sera with recombinant TS during 15 min at room temperature. Recombinant TS was expressed in a pTrcHis vector (Invitrogen, San Diego, CA) and purified by immobilized metal ion-affinity chromatography (HiTrap; GE Healthcare) according to the manufacturer's instructions (Buschiazzo et al. 1996). Remnant TS activity was determined by measuring the transference of sialic acid from 1 mM sialyllactose (1 mM, Sigma, St. Louis, MO) to 12 μM [
Ethical statements
Capture and handling of mammals in Santiago del Estero and Chaco provinces were conducted following the protocol no. 04223 and no. TW-01-004 approved by Institutional Animal Care and Use Committee protocol at the University of Illinois at Urbana Champaign and the Ethical Committee “Dr. Carlos Barclay” (Buenos Aires), respectively.
Animal care and use is guided by the International Guiding Principles for Biomedical Research Involving Animals developed by the Council for International Organizations of Medical Sciences.
Statistical analysis
The agreement between XD and TIA was expressed as co-reactivity, co-positivity, and co-negativity. The association between XD and TIA outcomes was determined by the Fisher exact test whereas concordance was established by calculating the Cohen kappa index. The Youden index was used to choose an appropriate cutoff value (Fluss et al. 2005).
Results
Evaluation of TS-neutralizing antibodies in wild mammals
We analyzed by TIA serum samples from different wild mammal species (n=158) previously diagnosed by XD for T. cruzi infection. The percentage of inhibition obtained for each group of mammals is presented in Figure 1. The cutoff value was set at 50% of inhibition by calculating the maximum Youden index for TIA assay in samples from marsupials (J=0.98), edentates (J=0.97), and mustelids (J=1.0).
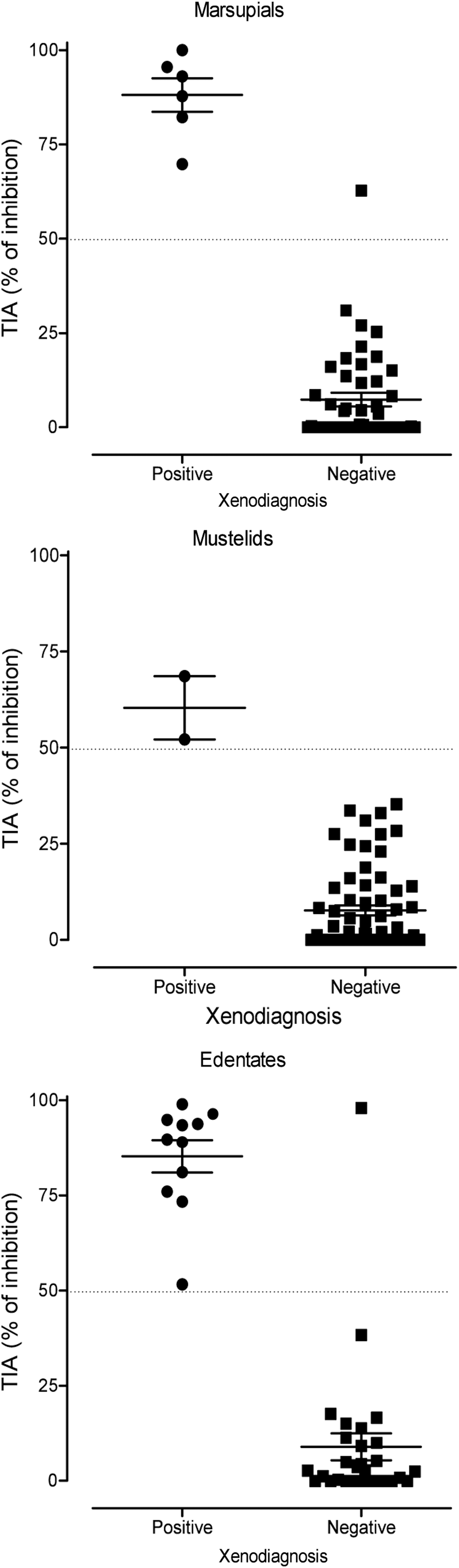
Trans-sialidase inhibition assay (TIA) reactivity in serum samples from sylvatic mammalian hosts that were analyzed by xenodiagnosis. The cutoff value is indicated by the dotted line. The Youden index was used to estimate the cutoff value.
The TIA mean value for XD-positive and XD-negative marsupials was 84.79±14.17 (n=5) and 5.91±8.67 (n=47), respectively; for mustelids, 60.36±11.57 (n=2) and 7.66±10.58 (n=64); and for edentates, 85.71±12.91 (n=11) and 5.74±8.55 (n=29), respectively.
The overall TIA versus XD co-reactivity was 98.7% (156/158). Co-positivity was 100% (18/18), whereas co-negativity was 98.5% (138/140). The comparative XD and TIA results for each mammalian species are shown in Table 1. All samples from XD-positive mammals were detected by TIA. One sample from a XD-negative marsupial (D. albiventris) showed a percentage of inhibition of 62.8% and one sample from a XD-negative edentate (D. novemcinctus) was also reactive by TIA (98.04%) (Table 1).
TIA and XD outcomes were significantly associated (p<0.0001), and showed a high degree of concordance between them (kappa index=0.89). The positive predictive value of TIA was 0.98 (95% confidence interval [CI] 0.92–0.99) and the negative predictive value was 1.0 (95% CI 0.82–1.0).
Discussion
Our study first describes that trans-sialidase neutralizing antibodies are elicited by different species of edentates, marsupials, and mustelids naturally infected. T. cruzi is classified in six discrete typing units (DTUs) that are differentially distributed among sylvatic, peridomestic, and domestic environments (Zingales et al. 2009). We have reported TS-neutralizing antibodies in mice experimentally infected with TcI (Risso et al. 2004), and in patients (Leguizamón et al. 1994, Leguizamón et al. 1998, Buchovsky et al. 2001, Blejer et al. 2008) and domestic dogs and cats (Sartor et al. 2011) from the southern cone of South America where TcII, TcV, and TcVI are the principal DTUs involved in domestic transmission cycles (Risso et al. 2004, Cardinal et al. 2008). In the current study, sera from two mustelids (Conepatus chinga) and 11 edentates (D. novemcinctus) infected by TcIII and five marsupials (D. albiventris) infected by TcI were reactive by TIA (Ceballos et al. 2006, Alvarado-Otegui et al. 2012). These findings demonstrate that TS-neutralizing antibodies are elicited during infections caused by different T. cruzi DTUs, showing the broad applicability of TIA to studies throughout endemic areas.
TIA was able to detect 100% of XD-positive samples from wild mammalian hosts. The co-negativity value was as high as 98.5% because TIA allowed the detection of two positive samples that had been XD-negative, a marsupial (D. albiventris) and an edentate (D. novemcinctus). The rare discrepancies recorded between XD and TIA values were most likely due to expected false-negative results by XD. The amplification by PCR of the variable region of T. cruzi kinetoplastid DNA (kPCR), and nuclear satellite (SAT-DNA-PCR) observed in samples from D. novemcinctus supports this conclusion (Orozco et al., unpublished results). The sensitivity of XD in seropositive dogs and cats naturally infected with T. cruzi ranged from 70% to 85% (Gürtler et al. 1996, Gürtler et al. 2007) and is influenced by parasitemia levels in a variety of wild mammalian hosts (Yabsley and Noblet 2002, Xavier et al. 2007, Roque et al. 2008).
Leishmania spp. do not express TS activity (Frasch 2000), and samples from patients or dogs suffering leishmaniosis were not TIA-reactive (Buchovsky et al. 2001, Sartor et al. 2011). It is highly likely that TIA may also be used to differentiate T. cruzi from Leishmania spp infections in wild mammals such as opossums, rodents, and edentates—all putative sylvatic reservoir hosts of L. infantum (Quinnell and Courtenay 2009).
TIA can be applied in serum samples obtained in field surveys to analyze and understand the persistence to T. cruzi in wild mammals. This method represents a novel tool for seroepidemiological studies and does not require the use of anti-immunoglobulins, thus solving a limitation of serological methods.
Footnotes
Acknowledgments
We are grateful to Julián Alvarado-Otegui, I. Colaianni, and G. Enriquez for technical assistance. M.S.L., R.E.G., and M.V.C. are Researchers and P.A.S. and M.M.O. are Fellows from Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina. Parts of this study were supported by awards from the National Institutes of Health/National Science Foundation Ecology of Infectious Disease program award R01 TW05836 funded by the Fogarty International Center and the National Institute of Environmental Health Sciences (to Uriel Kitron and R.E.G.), International Development Research Center (Ecohealth Program), University of Buenos Aires (to R.E.G. and M.S.L.), and National Agency of Science and Technology (to M.S.L.).
Author Disclosure Statement
No competing financial interests exist.