
Abstract
Bird migration is a recurring annual and seasonal event undertaken by more than 100 species of birds in the southeast Asian and northeast Palearctic regions that pass through or remain for short periods from April to May and September to November at Hong-do Island, Republic of Korea (ROK). A total of 212 ticks (40 Haemaphysalis flava, 12 H. longicornis, 146 Ixodes turdus, 13 I. nipponensis, and 1 I. ornithophila) were collected from 65/2,161 (3.0%) migratory birds consisting of 21 species that were captured from January, 2008, through December, 2009, as part of the Migratory Birds Center, Hong-do bird banding program for studying bird migration patterns. Adult ticks were assayed individually while larvae and nymphs were pooled (1–22 and 1–6 ticks per pool, respectively) into 31 and 65 pools, respectively. Ticks were assayed for zoonotic pathogens by PCR using 16S rRNA, heat shock protein (groEL), and internal transcribed spacer (ITS) gene primers to amplify genera specific for Anapalsma, Bartonella, and Borrelia PCR amplicons. Using the 16S rRNA-based nested PCR, A. phagocytophilum (n=1) was detected in I. nipponensis collected from Zoothera sibirica and A. bovis (n=1) was detected in I. turdus collected from Emberiza chrysophrys. Borrelia turdi 16S rRNA genes (n=3) were detected in I. turdus and I. nipponensis collected from Turdus pallidus and Zoothera aurea. Borrelia spp. 16S rRNA genes (n=4) were detected in Ixodes ticks collected from Emberiza tristrami, T. pallidus, and Z. aurea. The Bartonella grahamii ITS gene (n=1) was detected by nested PCR assay in I. turdus collected from Z. aurea. These results provide insight into the potential role of migratory birds in the dispersal of ticks and associated tick-borne pathogens throughout their ranges in Asia.
Introduction
Anaplasma phagocytophilum, the causative agent of human granulocytic anaplasmosis (HGA), transmitted primarily by Ixodes spp. ticks, has a northern hemisphere distribution (Chen et al. 1994). Recently in the Republic of Korea (ROK), A. phagocytophilum was detected by PCR technologies in deer and small mammals and in Haemaphysalis and Ixodes spp. collected by tick drag (Kim et al. 2006, Oh et al. 2009, Kang et al. 2011). Although Anaplasma bovis has been primarily detected in African cattle, it also was detected in Ixodes and Haemaphysalis spp. ticks, and deer in Japan and the ROK (Kawahara et al. 2006, Kim et al. 2006, Lee et al. 2009, Oh et al. 2009, Kang et al. 2011).
Bartonella grahamii, the zoonotic agent causing neuroretinitis, was first identified from a rodent in the United Kingdom (Birtles et al. 1994, Kerkhoff et al. 1999). In Asia, B. grahamii is the predominant species detected from rodents in neighboring countries, i.e., China (Liu et al. 2010) and Japan (Inoue et al. 2009, Kabeya et al. 2011).
Borrelia turdi was firstly characterized and isolated from Ixodes turdus, a less commonly collected species from Japan (Fukunaga et al. 1996), and more recently from Ixodes ricinus collected from passerine birds from Norway (Hasle et al. 2011). Both papers indicated that the host of B. turdi was birds, but the pathogenesis of B. turdi is not known and reports have been rare.
The purpose of this investigation was to determine the prevalence of Anaplasma, Bartonella, and Borrelia species in ticks collected from migratory birds that are potential hosts for the introduction of exotic tick species and reservoirs for the dispersal of the pathogens throughout their migratory routes in southeast Asia.
Materials and Methods
Survey area
The Migratory Birds Center of the National Park Research Institute, located on Hong-do Island (34°41’07”N, 125°11’33”E), conducted bird banding surveys from January, 2008, to December, 2009, to understand migration routes and strategies of migratory birds (Fig. 1). Hong-do Island, designated as National Monument No. 170 (April, 1965), is a small (6.4 km2) protected remote southwestern island located in Heuksan-myeon (district), Shinan-gun (county), Jeollanam-do (province), and 115 km west of Mokpo (mainland port city). Migratory birds pass through Hong-do and the ROK mainland during the spring and autumn migration along their south and northeast Asian flyways.
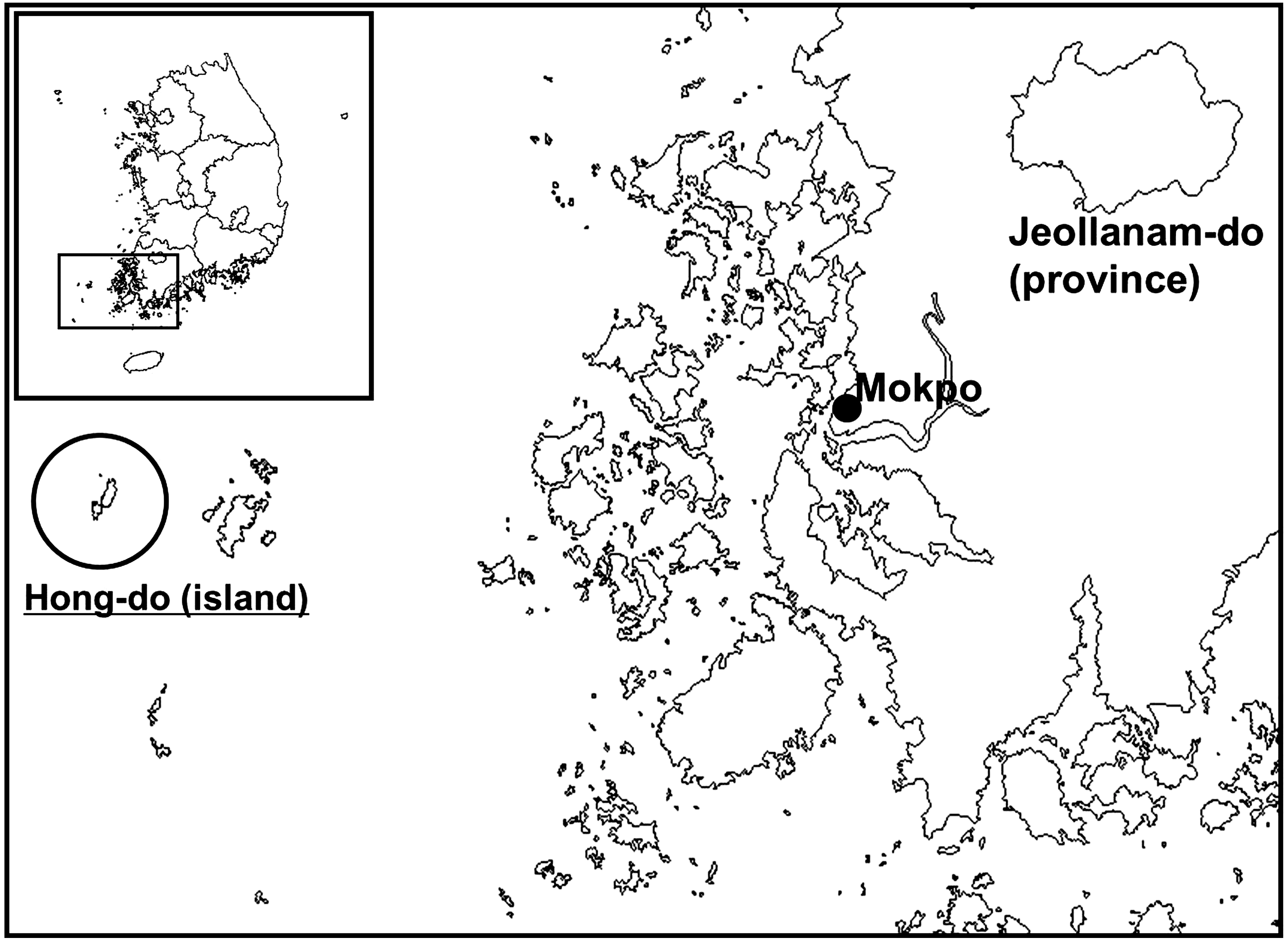
Hong-do Island, the collection site of ticks removed from migratory during the Migratory Birds Center bird banding surveys, Jeollanam-do Province, Republic of Korea.
Bird and tick collections
Migratory birds were captured on Hong-do using two 36-mm-meshed mist nets (12 meters in length, 2.5 meters high) placed at the ground level. The trapped birds were identified to species, banded with an identification number, and their sexes and ages determined. Banded birds were examined closely, particularly around the head and neck, for ticks and other ectoparasites prior to release. Ticks were secured by their mouthparts next to the skin with a fine forceps and gently removed to prevent injury to the bird. The removed ticks were placed individually in cryovials containing 70% ethyl alcohol and labeled with a unique identification number that corresponded to the migratory bird collection data. The ticks were identified microscopically to stage of development and species using taxonomic identification keys (Yamaguti et al. 1971, Hoogstraal and Wassef 1973). An electronic data sheet, which included the unique bird identification number, species, sex, and other pertinent information, was completed, and the ticks were assayed by PCR techniques for rickettsial and bacterial pathogens.
DNA extraction and PCR amplification
Ticks were pooled by tick species, life stages, sex (adults), collection dates, and unique bird collection numbers for each species. Ticks were homogenized mechanically using a Beadbeater TissueLyser II (QIAGEN) with 180 μL of lysis buffer, 20 μL of proteinase K (600 AU/mL), and 5-mm stainless steel beads at 30 frequencies/s for 5 min, followed by incubation at 56°C overnight and then centrifugation at 12,000×g for 10 min at room temperature. After centrifugation, the supernatant was used for genomic DNA extraction performed with DNeasy® Tissue Kits (QIAGEN) according to the manufacturer's instructions.
PCR and nested PCR were performed using specific primers for A. phagocytophilum, A. bovis, and Bartonella and Borrelia spp. (Table 1). A. phagocytophilum genomic DNA, provided by J. Stephen Dumler (Johns Hopkins University School of Medicine, Baltimore, MD), was used as a positive control for detecting Anaplasma species. Bartonella henselae and B. burgdorferi isolates purchased via the American Type Culture Collection were used as positive controls. The first and nested PCRs were performed in a total volume of 25 μL. Each PCR mixture consisted of 10 pmol of primers, 1 U recombinant Taq DNA polymerase (Takara Bio, Inc.), 10×PCR buffer (Takara Bio, Inc.), 2.5 mM deoxyribonucleotide triphosphates (dNTPs) mixture (Takara Bio, Inc.), 10- to 100-ng samples of genomic DNA for the first PCR, and 1 μL of the first PCR product for the second PCR. The amplification of PCR products was carried out in a PTC-200 thermal cycler (MJ Research, Inc.) as described in Table 1.
Cloning, nucleotide sequencing, and phylogenetic analysis
The PCR products were purified with QIAquick Gel Extraction kits (QIAGEN). After purification, the amplicons were cloned with pGEM®-T Easy Vectors (Promega) followed by transformation into Escherichia coli DH5α, and then plated onto LB agar containing 50 μg/mL of ampicillin. Plasmid DNA for sequencing was purified using the Wizard® Plus SV Minipreps DNA Purification System (Promega) according to the manufacturer's instructions. Purified recombinant plasmid DNA was sequenced using a T7 and SP6 promoter primer set by dideoxy termination with an automatic sequencer (ABI 3730xl capillary DNA sequencer, Applied Biosystems). The obtained sequences were evaluated with Chromas software (Ver 2.33,
Nucleotide sequence accession numbers
The GenBank accession numbers of 16S ribosomal (r)RNA, groEL, and internal transcribed space (ITS) gene sequences and specific genospecies sequences related to bacterial pathogens for sequence comparisons are included in Figures 3 –6, below.
Results
A total of 3,816 birds representing 102 species were banded and examined for ectoparasites from January, 2008, to December, 2009 (Fig. 2), and 212 ticks were removed from 65 birds belonging to 21 species and 12 genera (Table 2). The majority of ticks were collected from White's Thrush (Zoothera aurea) (114 ticks, 53.8%), followed by the Pale Thrush (Turdus pallidus) (27 ticks, 12.7%), the Yellow-throated Bunting (Emberiza elegans) (20 ticks, 9.4%), and Naumann's Thrush (Turdus naumanni) (10 ticks, 4.7%). The remaining 17 species accounted for 19.3% (ranging from 1 to 6 ticks, 0.5–2.8%) of the total number of ticks collected. A total of 5 species (larvae, nymphs, and adults) of ticks, belonging to 2 genera, were identified (Tables 1 and 3). The most frequently collected tick was I. turdus Nakatsuji (146, 68.9%), followed by Haemaphysalis flava Neumann (40, 18.9%), Ixodes nipponensis Kitaoka and Saito (13, 6.1%), Haemaphysalis longicornis Neumann (12, 5.7%), and Haemaphysalis ornithophila Saito, Hoogstraal, and Wassef (1, 0.5%) (Table 1).

Monthly distribution of total number of captured and banded migratory birds, and number of ticks, by species, collected from the migratory birds at Hong-do Island from January, 2008, through December, 2009, Jeollanam-do Province, Republic of Korea.
PCR-positive pathogens were determined by the maximum likelihood estimates (MLE), based on the number of positive individuals/pools and pool size for ticks tested.
A. phagocytophilum and A. bovis 16S rRNA genes were identified by species-specific nested PCR. A. phagocytophilum was detected in I. nipponensis collected from Zoothera sibirica and A. bovis was detected in I. turdus collected from Emberiza chrysophrys (Table 3). The genome sequences were analyzed and compared with the fragments of 16S rRNA gene sequences to demonstrate genetic relationships between Anaplasma spp. detected in ticks. The acquired A. phagocytophilum 16S rRNA gene sequence (JX219480) corresponded to previously sequenced A. phagocytophilum (GU556624) from the ROK (Fig. 3A), whereas the acquired A. bovis 16S rRNA gene sequence (JX219481) was 99.8% similar to A. bovis sequence (GU556626) from the ROK (Fig. 4).

(
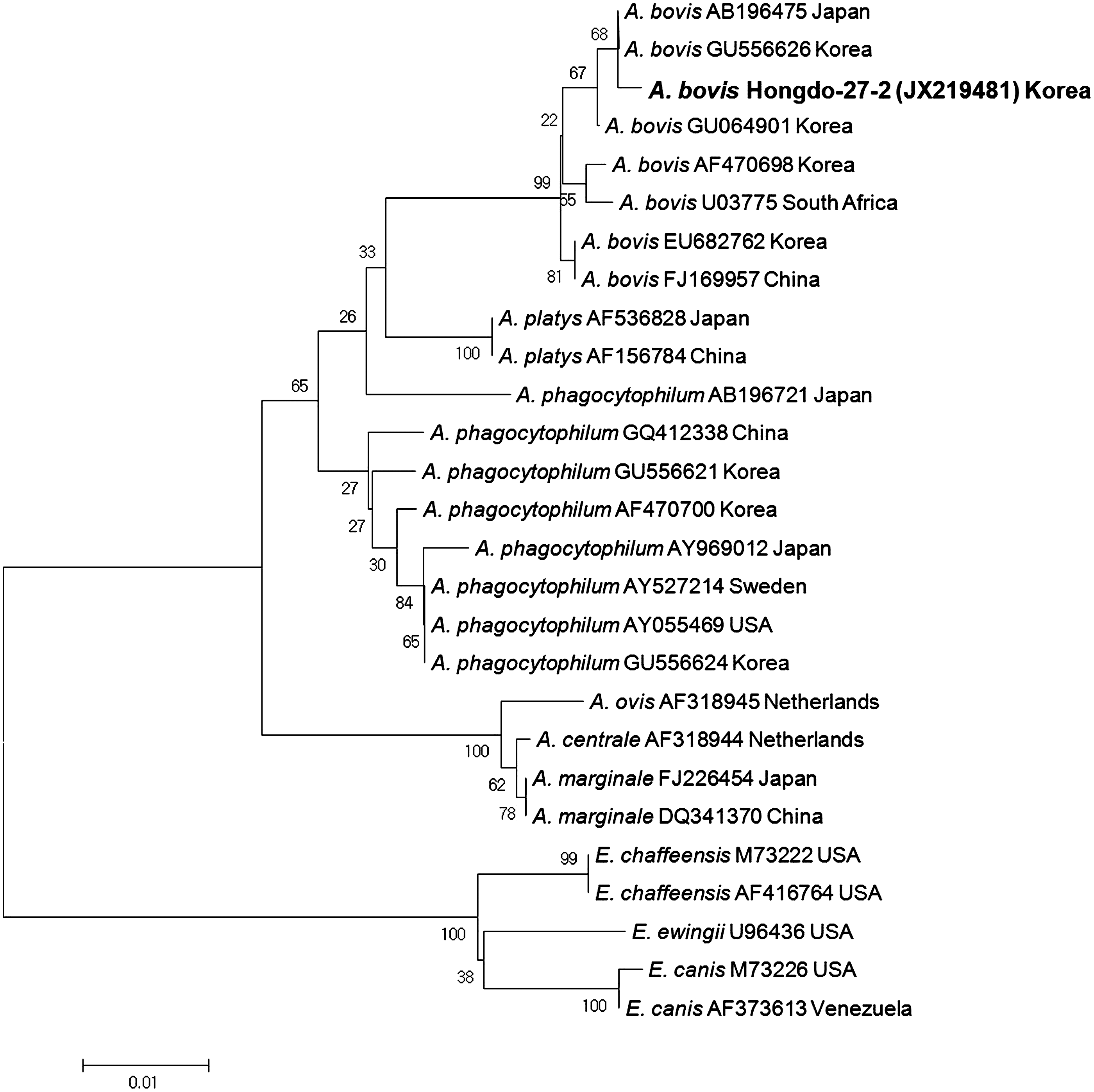
Phylogenetic relationships among Anaplasma bovis (boldface) detected in ticks collected from migratory birds in the Republic of Korea and Anaplasma and Ehrlichia species based on partial nucleotide sequences of the 547-bp 16S rRNA genes. The neighbor-joining method was used to construct a phylogenetic tree. The numbers at nodes are the proportion of 1,000 bootstrap iterations that support the topology shown.
Using the groEL gene-specific primer pairs shown in Table 1 for PCR, one A. phagocytophilum groEL gene nucleotide sequence (JX219474) was obtained. The product size was 1,297 base pairs (bp) and translated 409 amino acids of groEL. The obtained groEL amino acid sequence was identical to A. phagocytophilum (ADO34908) from the ROK (Fig. 3B).
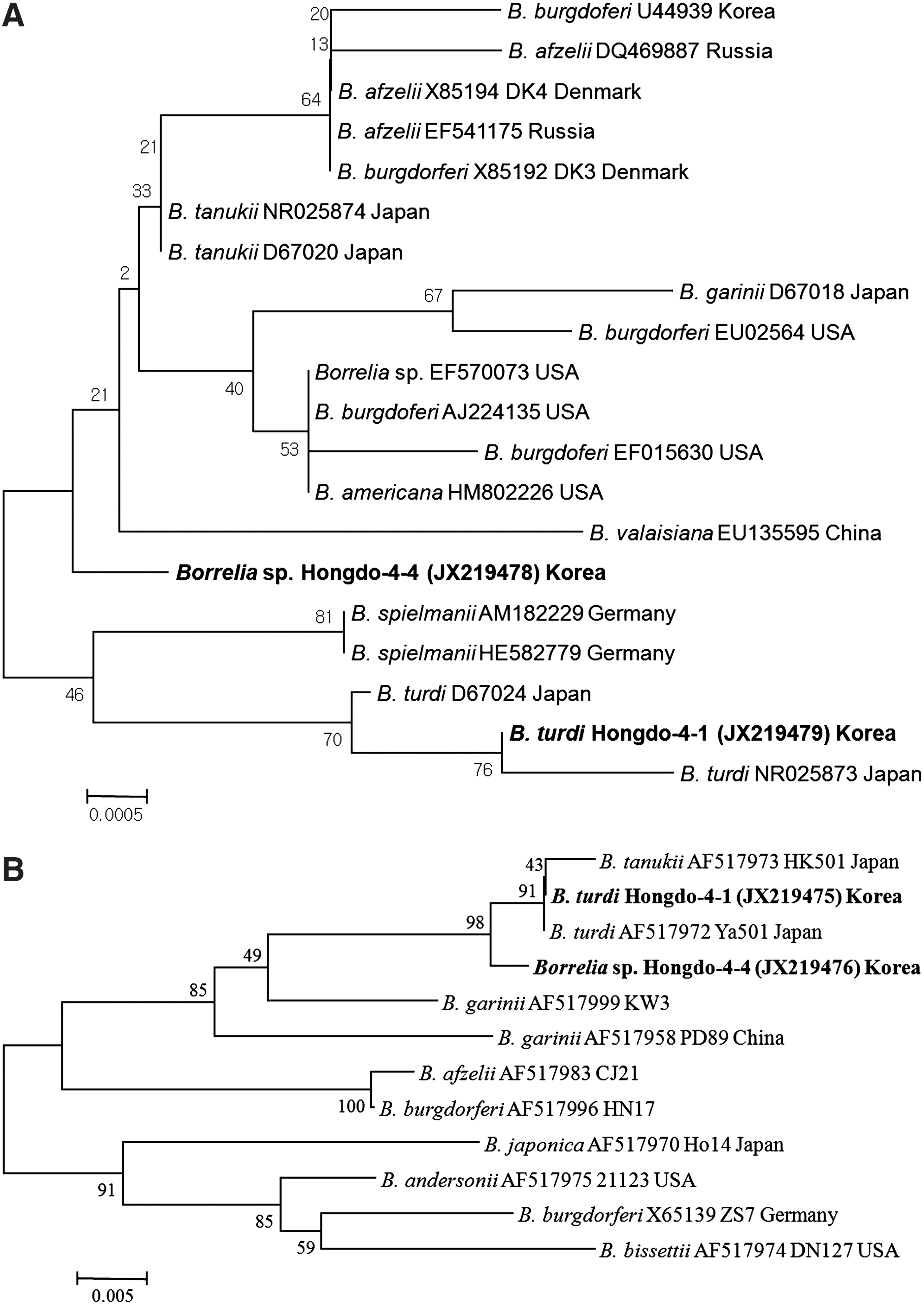
Borrelia spp. (n=4) and B. turdi (n=3) 16S rRNA genes were identified by nested PCR assay. B. turdi were detected in 2 I. turdus and 1 I. nipponensis collected from Z. aurea (2) and T. pallidus (1), whereas Borrelia spp. were detected in 3 I. turdus and 1 I. nipponensis collected from Emberiza tristrami (1), T. pallidus (2), and Z. aurea (1). The genetic relationships between 7 genome sequences of Borrelia spp. detected in ticks were analyzed and compared with the fragments of 16S rRNA gene sequences (Fig. 5A). The Borrelia spp. 16S rRNA gene sequences (JX219478) were identical and showed 99.9% homology to Borrelia tanukii (NR025874) from Japan. The obtained B. turdi 16S rRNA gene sequences (JX219479) were identical and showed 99.9% homology to B. turdi (NR025873 and D67024) from Japan (Fig. 6).
(
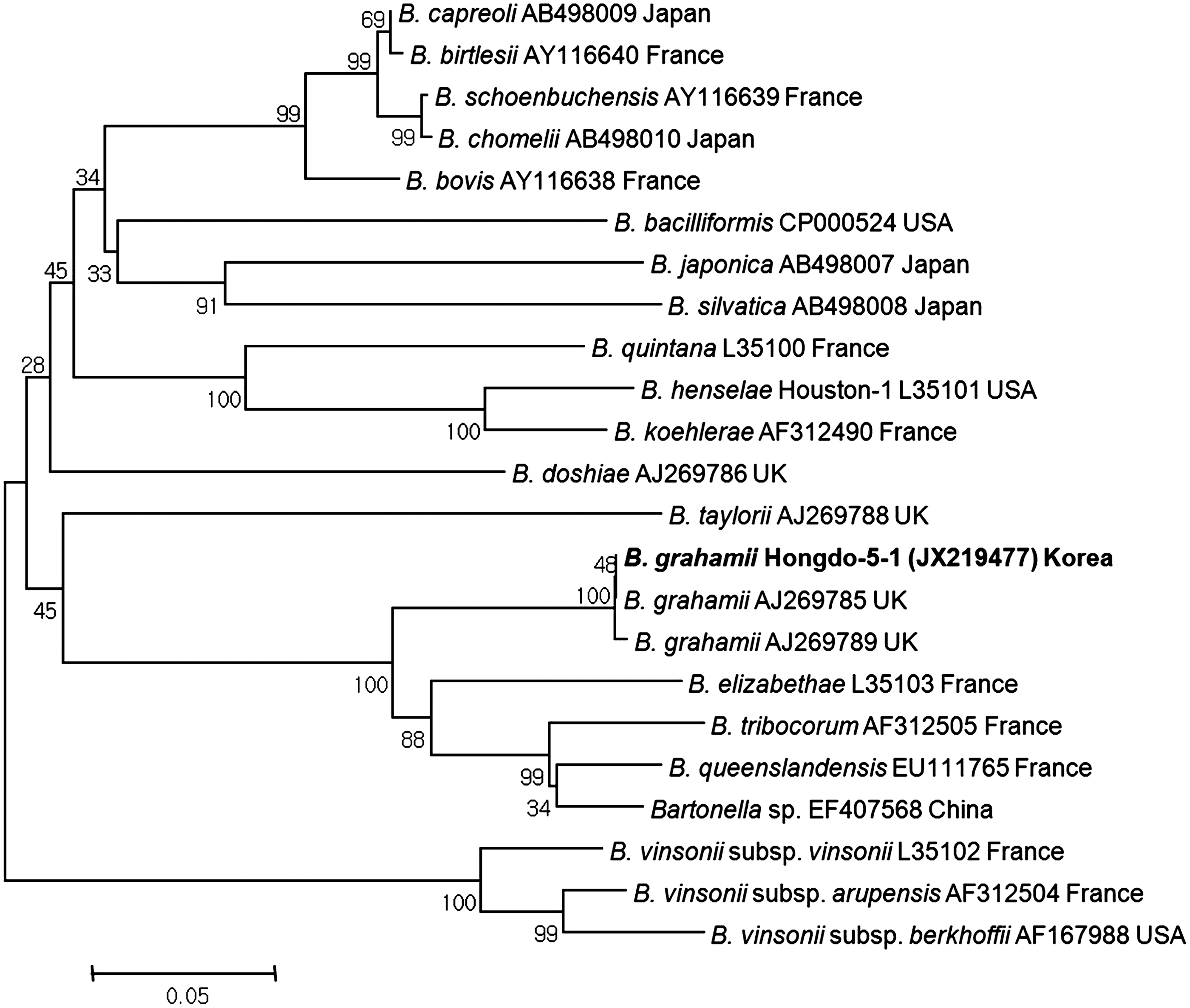
Phylogenetic relationships among Bartonella grahamii (boldface) detected in ticks collected from migratory birds and Bartonella species based on partial nucleotide sequences of the 484-bp internal transcribed spacer (ITS) genes. The neighbor-joining method was used to construct a phylogenetic tree. The numbers at nodes are the proportion of 1,000 bootstrap iterations that support the topology shown.
Using the groEL specific nested PCR, 2 Borrelia spp. (JX219476) and 2 B. turdi (JX219475) groEL gene sequences were detected in ticks collected from migratory birds. The product size was 310 bp and contained coding sequence for 103 amino acids of groEL. In spite of being characterized as different species, 99.4% homology was shared between the 2 Borrelia spp. and 2 B. turdi groEL genetic sequences (Fig. 5B), whereas all of the amino acid sequences were identical. The Borrelia spp. and B. turdi groEL genes were different at 2 sites (nucleotide positions 113 and 205) of 310 nucleotide sequences. The 2 Borrelia spp. groEL gene sequences were identical and showed 99.4% and 99.0% homology to B. turdi (AF517972) and B. tanukii (AF517973), respectively. Two B. turdi groEL gene sequences were identical each other and showed 100.0% and 99.7% homology to B. turdi (AF517972) and B. tanukii (AF517973), respectively (Fig. 5B).
The B. grahamii ITS gene (JX219477) was detected by nested PCR assay in I. turdus collected from Z. aurea (Table 3). The product size was 484 bp and contained noncoding sequences. To investigate the genetic relationship between the Bartonella species detected in ticks, the ITS sequence was analyzed and compared with the fragments of ITS gene sequences. The acquired B. grahamii ITS gene sequence was identical to B. grahamii (AJ269785) in UK (Fig. 6).
Discussion
The bird population of Korea is augmented by millions of birds that migrate or stop over from April to May and September to November. More than 100 species of migratory birds are recorded annually on their spring migrations to their summer breeding grounds in the northeastern Palearctic region, including Russia and eastern China, and autumn migrations to their nonbreeding grounds in southeast Asia and Australia (Pospelova-Shtrom et al. 1965, Filippova 1984, Lee et al. 2003). These bird migrations are behavioral responses, i.e., traversing long distances to their breeding and overwintering grounds that are affected by changes in food availability, habitat, and weather. Emberiza, Zoothera, and Turdus spp. migrate annually and breed in the Siberian taiga zone during the summer and return to southeast Asia during the winter. Turdus spp. are known to have relatively high tick infestation rates due to their habit of spending a majority of their time on the ground in search of food among grasses and low-lying vegetation where ticks are commonly found (Ishiguro et al. 2000, Hasle et al. 2009a, b).
I. turdus is primarily an ectoparasite of birds (Yamauchi et al. 2001). During this surveillance period, it was collected from 12 of the 21 bird species identified with ticks and was the most frequently collected tick species (66.2% of all specimens). These results are similar to migratory bird surveys conducted in the eastern Shimane Prefecture, Japan, where I. turdus accounted for 79.0% of all ticks collected from birds (Yamauchi and Mori 2004). Haemaphysalis ornithophila is widespread in southeast Asia and China, but is infrequently collected from birds throughout its range. Only 1 specimen was collected from a White's Thrush (Z. aurea) during this survey, whereas 2 adults were collected in previous bird banding surveys in the ROK (Kim et al. 2009). Z. aurea may transport this tick, in addition to other species, when migrating to their summer breeding grounds in Russia (Kim et al. 2009).
A. phagocytophilum and A. bovis are obligate intracellular tick-borne bacteria in the family Anaplasmataceae with Ixodes spp. as the primary vectors. Limited studies of birds as reservoirs of A. phagocytophilum have been conducted. A. phagocytophilum DNA was detected in I. ricinus and I. scapularis collected from migratory birds in Canada, Norway, Poland, Russia, Spain, and Sweden (Alekseev et al. 2001, Bjöersdorff et al. 2001, Skotarczak et al. 2006, Ogden et al. 2008, Paulauskas et al. 2009, Hildebrandt et al. 2010, Palomar et al. 2012). These studies presented a low prevalence of A. phagocytophilum in ticks collected from migratory birds, suggesting that birds were not a competent reservoir for A. phagocytophilum. In this study, only 1 A. phagocytophilum and 1 A. bovis were detected in I. nipponensis and I. turdus, respectively. The low prevalence of Anaplasma spp. in our study was similar to previous reports (Alekseev et al. 2001, Bjöersdorff et al. 2001, Skotarczak et al. 2006, Ogden et al. 2008, Paulauskas et al. 2009, Hildebrandt et al. 2010, Palomar et al. 2012). Nevertheless, migratory birds might be an important component in the dispersal of ticks infected with endemic Anaplasma spp. throughout their range where they overwinter and breed.
B. turdi has not been reported to cause disease in humans and was previously only detected in I. turdus ticks collected from migratory birds in Japan (Fukunaga et al. 1996). Recently in Norway, Hasle et al. (2011) detected B. turdi in I. ricinus collected from migratory birds, but failed to show the gene sequences and phylogenetic analysis. In this study, Borrelia spp. sequences in I. nipponensis and I. turdus collected from migratory birds were divergent from previously reported Borrelia spp. sequences (Fig. 5). The Hongdo-4-1 genotype (JX219475) belonged to the B. turdi clade, but was divergent from other reported B. turdi sequences. The Hongdo-4-4 genotype (JX219476) did not separate into any of the Borrelia spp. subclades. These results and reported studies suggest that the vector of B. turdi is I. turdus and the reservoirs are ground-dwelling birds.
Fleas have been identified as the vectors for many Bartonella spp. The rodent flea (Ctenocephalides nobilis nobilis) has been identified as the primary vector of B. grahamii (Bown et al. 2004). More recently a number of tick species have been shown to be positive for Bartonella spp. based primarily on PCR techniques or infrequently by culture (Billeter et al. 2008). In the ROK, Bartonella DNA has been detected in H. flava, H. longicorinis, I. nipponensis, and I. turdus ticks collected from rodents or by tick drag (Kim et al. 2005). In this study, the B. grahamii ITS gene sequence (JX219477) was detected in I. turdus ticks collected from the Pale Thrush (T. pallidus). However, there were insufficient ITS gene sequences available for a comprehensive comparison with sequences reported in GenBank. Additional phylogenetic analysis for gltA, groEL, and rpoB genes targeting nested PCR was conducted, but unfortunately a positive band was not obtained. Nevertheless, this result indicates that I. turdus and migratory birds may be potential vectors and reservoirs of Bartonella spp. in Asia.
Hard ticks are the primary vectors of a variety of bacterial pathogens, including Anaplasma, Borrelia, and Ehrlichia spp. (Parola et al. 2001). Although Bartonella spp. are transmitted by other ectoparasitic arthropods, the only evidence to support the possibility of tick-borne transmission is indirect (Angelakis et al. 2010). Recently, because of the rapidly expanding number of reservoir-adapted pathogens that have been discovered, efforts to clarify the vector competence, potential for transmission, and disease health risks are relevant to public and veterinary health. Herein, we present the various tick-borne pathogens, i.e., Anaplasma, Bartonella, and Borrelia spp. that were detected in Ixodes spp. from migratory birds. These results do not conclusively identify the role of migratory birds as reservoirs or introduction of exotic ticks in nonendemic areas, but they provide an insight into the potential role of migratory birds in the dispersal and infectious cycles of tick-borne pathogens in Asia.
Footnotes
Acknowledgments
This research was supported by a National Research Foundation of Korea Grant funded by the Korean Government (2011-0015349), and funding for portions of this work was provided by the Armed Forces Health Surveillance Center, Global Emerging Infections Surveillance and Response System, Silver Spring, MD, and the National Center for Military Intelligence, Ft. Detrick, MD. We thank the members of Migratory Bird Center, National Park Research Institute, for bird tick collections during their bird-banding surveys. We especially thank Dr. Joel Gaydos, Global Emerging Infections Surveillance and Response System, Silver Spring, MD, USA, for his support and constructive criticism.
Author Disclosure Statement
The opinions expressed in this article are those of the authors and do not reflect official policy or positions of the US Department of the Army, the US Department of Defense, or the US Government.