
Abstract
Following the West Nile Virus (WNV) epidemic in 2010 in Central Macedonia, Greece, which resulted in 197 human neuroinvasive disease cases, we determined the seasonal appearance and prevalence of the virus in 2011 by testing weekly for WNV genomic RNA in mosquitoes collected in carbon dioxide–baited traps, and for anti-WNV antibodies in sentinel chickens. Preliminary findings of the surveillance program regarding the circulation of “Nea Santa-Greece-2010” in sentinel chickens were rapidly communicated to public health authorities. In the present article, the full 2011 data produced by this surveillance program are presented. We detected enzootic circulation of WNV in chickens 1 month prior to the onset of the first human cases in 2011. Culex pipiens and Cx. modestus were abundant throughout the sampling period and at all sites of increased transmission. Molecular identification and phylogenetic analysis of WNV isolates from two chickens and one Cx. pipiens mosquito pool suggested that: (1) the virulent “Nea Santa-Greece-2010” WNV lineage 2 strain responsible for the 2010 epidemic was actively circulating in 2011, and (2) all Greek isolates belong to a distinct recent evolutionary clade. In Europe, where numerous strains of different virulence coexist, sequencing information for WNV is important for phylogeography and identification of virulent strains for human health risk assessment.
Introduction
WNV strains belonging to lineages 1 and 2 have varying virulence and neuroinvasiveness properties (Beasley et al. 2002). Neuroinvasive isolates mainly belonged to lineage 1, whereas lineage 2 strains were considered to be less virulent (Venter and Swanepoel 2010). Before 2010, WNV epidemics in Europe were caused mainly by linage 1 strains. However, a neuroinvasive lineage 2 strain was responsible for a major WNV outbreak in 2010 in Greece (Region of Central Macedonia) with 197 human neuroinvasive disease [West Nile neuroinvasive disease (WNND)] cases and 35 fatalities (Danis et al. 2011). A new epidemic occurred in 2011 with 76 WNND cases and eight fatalities (Hellenic Center for Disease Control and Prevention 2011). During the 2010 epidemic, the Greek lineage 2 strain (“Nea Santa-Greece-2010”) was detected in Culex mosquitoes and in donated human blood (Papa et al. 2012), as well as in an infected Eurasian magpie (Valiakos et al. 2011). The “Nea Santa-Greece-2010” virus was highly similar (99.6%) to the goshawk-Hungary-2004 WNV strain circulating in Hungary from 2004, and later on detected in Austria (2008) and Italy (2011) (Bakonyi et al. 2006, Wodak et al. 2011, Savini et al. 2012). Interestingly, despite the close genetic relation to the Austrian and Hungarian lineage 2 viruses, the Greek strain presented high virulence in mammals, as evidenced by the high number of neuroinvasive disease cases in humans. An amino acid substitution (H249P), absent from the closely related Hungarian and Italian lineage 2 strains, is suspected to be responsible for the high neuroinvasiveness and virus circulation observed during the Greek epidemic (Papa et al. 2011). Previous experimental studies have attributed this mutation to a higher capacity of the virus to replicate in American corvids, which likely would result in increased virus transmission rates (Brault et al. 2007).
WNV arrived in the United States in 1999, caused large outbreaks, and rapidly spread throughout most of the country by the end of 2002 (Centers for Disease Prevention and Control 2003). Unlike the situation in Europe, virus strains circulating in the United States belong solely to WNV lineage 1 (Kramer at al. 2008). Upon arrival of WNV, a variety of arbovirus surveillance systems were established in the United States. These systems included avian-, mosquito-, and nonhuman mammal-based sampling to facilitate prediction and prevention of human and domestic animal infection (Centers for Disease Control and Prevention 2003). Similar arbovirus surveillance systems have been used in Europe, with limited reports on the efficiency, sensitivity, and early reporting capacity of active bird monitoring and mosquito screening (Cernescu et al. 2000, Rizzoli et al. 2007, Calistri et al. 2010b). While many studies in the United States and Europe have addressed surveillance methodology for lineage 1 WNV infection, little is known about the appropriate surveillance schemes in the context of lineage 2 WNV circulation. Furthermore, due to the presence of multiple strains of variable virulence within Europe, it is important to adapt an arbovirus surveillance strategy that not only allows for early detection of enzootic transmission, but also for molecular characterization of the circulating viruses. Such arbovirus surveillance may be necessary for better assessment of public health risk and guidance for vector management interventions in Europe.
As a result of the 2010 epidemic in Greece, a vector and sentinel chicken surveillance program was initiated in 2011. Preliminary findings of that program were rapidly communicated to public health authorities (Chaskopoulou et al. 2011a). More specifically, active circulation of the “Nea Santa-Greece-2010” strain was detected early in the epidemic season in one seroconverted sentinel chicken, indicating that the virus may have overwintered in northern Greece.
The present article compiles the full 2011 data produced by the surveillance program, which specifically (1) provided weekly monitoring for WNV in sentinel chickens and mosquitoes around Thessaloniki, Greece, (2) used sentinel chickens as an early warning indicator for human WNV infection in the region, (3) isolated and molecularly characterized the virus in sentinel chickens and Cx. pipiens mosquitoes, and (4) investigated the activity patterns of the most prevalent Culex mosquito species that may be associated with pathogen transmission. These data, along with the WNND case onsets reported in the area, were discussed to assess the value and sustainability of this program as an early warning indicator for WNV epidemics in Europe.
Materials and Methods
Study Area—Regional Unit of Thessaloniki
The Regional Unit of Thessaloniki occupies the center of Central Macedonia Region in Northern Greece and has a population of ∼1.5 million people. The Regional Unit of Thessaloniki is divided into 14 municipalities, with the majority of the population aggregated in the urban municipalities (urban complex) of the city and the rest in the suburban and rural municipalities (Fig. 1). In terms of hydrology, the territory has significant natural wetlands, rivers, and ∼20,000 hectares of rice fields (European Center for Disease Control and Prevention 2010) with prolific mosquito breeding sites (Chaskopoulou et al. 2011b). The western river deltas are foci for migratory and native birds. Trapping and sentinel surveillance were conducted around the perimeter of the city's urban complex.
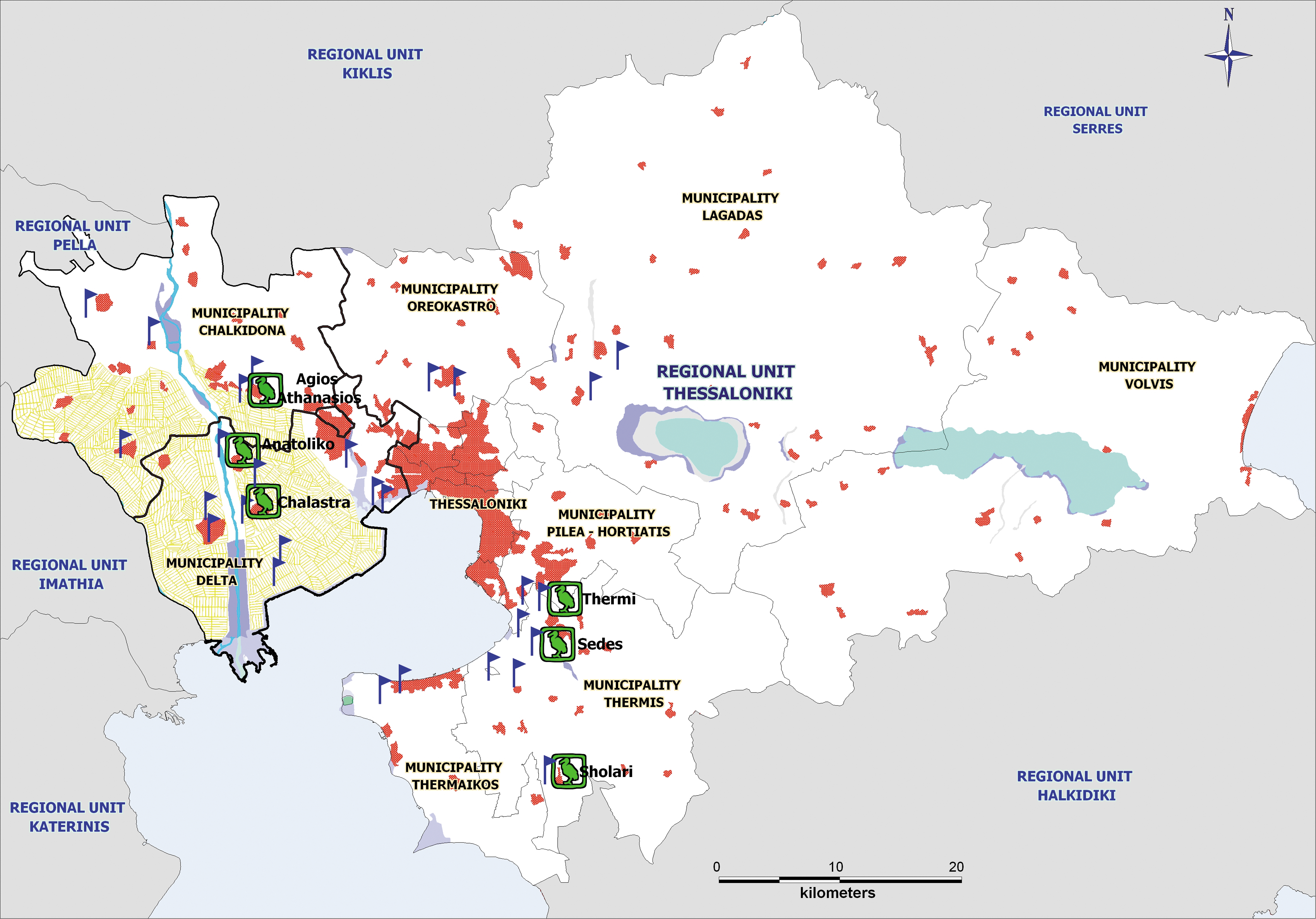
Regional unit of Thessaloniki. The map shows the locations of the 28 CDC light traps (blue flags) along the western, eastern and northern suburban/rural regions of the city, and the six sentinel chicken coops (poultry symbol, Agios Athanasios, Anatoliko, Chalastra, Thermi, Sedes, and Scholari). The topography of the region is depicted on the map (urban areas in red, water bodies/wetlands in blue, agricultural areas/rice-fields in yellow).
Sentinel chickens
Six cages, each one holding six chickens aged 2.5 months, were exposed from May to October, 2011—two at the Delta municipality, one at the Chalkidona municipality, and three at Thermi municipality. These municipalities had the highest incidences of human cases during the 2010 epidemic, high mosquito densities, and close proximity to the urban complex. The cages were placed within or in close proximity to residential communities that experienced abundant mosquito activity and were characterized by prolific mosquito breeding sites. Chicken plasma and sera were collected weekly. Upon seroconversion, chickens were replaced. Percent seroconversion (numbers of converted chickens/total number of chickens used) was calculated by municipality.
Mosquito surveillance
Carbon dioxide (dry ice)-baited CDC traps (John W. Hock, Gainesville, FL) were deployed weekly at 28 sites beginning May 20, 2011 (Fig. 1). Six traps co-localized with sentinel chicken flocks; the remaining traps were placed along the eastern, northern, and western outskirts of the city. Weekly mosquito catches were counted and species were identified (Darsie and Samanidou 1997, Schaffner et al. 2001). Catches >200 females per light trap were identified using a random sample of 200 mosquitoes, so that total species numbers could be extrapolated to counts for the entire sample. Only Culex population dynamics over time and temperature (recorded from ground weather station, Kestrel 4500 NV Weather Tracker; Nielsen-Kellerman Co., Boothwyn, PA) were followed. WNV detection was conducted using pools of 50 mosquitoes collected 1 and 2 weeks prior to chicken seroconversions (collected from traps adjacent to cages with chickens that seroconverted) and also from traps in the rice fields and Axios river regions because they are considered prolific mosquito breeding sites with high activity of migratory and native birds.
Laboratory testing of sentinel chickens and mosquitoes
Serum samples were assayed by competitive enzyme-linked immunosorbent assay (cELISA) for the detection of WNV-specific antibodies using a commercial kit (ID Screen West Nile Competition, IDVet, Montpellier, France) with modifications in incubation times to increase sensitivity of detection. Briefly, after addition of the samples to the wells, the plate was incubated overnight at 4°C, and after the conjugate addition the incubation was performed for 60 min at 26°C. WNV seropositivity was confirmed by microtiter plaque reduction neutralization test (micro-PRNT), according to Weingartl et al. (2003).
Mosquitoes were pooled into groups of 50 in 2-mL microcentrifuge tubes with two 4-mm glass balls and 1 mL of phosphate-buffered saline (PBS) containing 1% of inactivated fetal bovine serum (FBS; Sigma-Aldrich, Steinheim, Germany). Disruption was performed for 80 s at speed 4.0 using a RiboLyser homogenizer (Hybaid, Ltd., Teddington, UK). The homogenate was centrifuged (16,000×g, 5 min at 4°C) and the supernatant was stored at −70°C.
Plasma samples were stored at −70°C. RNA was extracted retrospectively from plasma obtained from seroconverted chickens using the QIAamp Viral RNA Mini Kit (Qiagen, Hilden, Germany). RNA isolation from the clarified mosquito homogenates was performed according to O'Guinn et al. (2004).
RNA extracts were examined using a one tube real-time RT-PCR protocol employing a primer pair (WNPolUp and WNPolDo2) and a hydrolysis probe (WNPolProb2) designed for the specific detection of WNV and targeting a 144-bp part of the nonstructural protein 5 (NS5) gene. Primers and probe were selected from highly conserved NS5 regions of lineage 1 and lineage 2 sequences available in the GenBank. Amplification reactions were run in a total volume of 25 μL using 5 μL of RNA extract and 20 μL of reaction buffer of a commercial RT-PCR kit (One Step RT-PCR, Qiagen, Hilden, Germany). The assay was performed using the CFX96 Touch™ Real-Time PCR detection system (Bio-Rad Laboratories, Hercules, CA). Amplification conditions and primers are described in Table 1. The detection range and analytical sensitivity of the real-time RT-PCR were evaluated using two lineage 1 strains (PaAn001/France and Eg101/Egypt) and the lineage 2 “Nea Santa-Greece-2010” virus. For this purpose, the 50% tissue culture infectious dose (TCID50)/mL of these strains was determined in Vero cells, using the Reed and Muench formula.
ZNA probe with 5 cationic spermine units purchased from Metabion International AG, Martinsried, Deutschland.
Samples found positive by the real-time RT-PCR screening protocol were confirmed using a two-step RT-PCR designed for the detection of WNV lineage 2 isolates. cDNA was synthesized by reverse transcription (RT), according to Dovas et al. (2010). Two microliters of cDNA were used in a PCR assay employing a primer pair (WN-NS3up2 and WN-NS3do2) targeting a 423-bp part of the nonstructural protein 3 (NS3) gene. Primers were selected from highly conserved NS3 regions of all lineage 2 sequences available in the GenBank database. The PCR amplification was performed in 50-μL reactions by using 1×PCR buffer, 0.2 mM of each dNTP, 4 units of HotStarTaq DNA polymerase (Qiagen, Hilden, Germany), and diethylpyrocarbonate (DEPC)-treated water. Cycling conditions and primers are described in Table 1.
The NS3 gene was sequenced in all samples confirmed as WNV positive. Three PCR assays amplifying overlapping fragments of the NS3 were applied using respective primer pairs: WNUp4579 and WNDo5273 (695 bp), WN-NS3up2 and WNDo5917 (702 bp), and WNUp5766 and WNDo6504 (739 bp). Cycling conditions and primers are described in Table 1. The PCR products were purified and sequenced. Phylogenetic analysis of WNV NS3 sequences was performed using MEGA 5.0 software (Tamura et al. 2011).
WNV isolation was performed in a biosafety level-3 (BSL-3) laboratory. Vero cells (ECACC 84113001; ECACC, Wiltshire, United Kingdom) were used, grown in Dulbecco's Modified Eagle's medium (DMEM; Invitrogen-Gibco, Groningen, The Netherlands) containing 10% FBS and antibiotics. Vero cell monolayers were inoculated with 10-fold serial dilutions of the RT-PCR–positive chicken plasma samples. After 1 h of incubation at 37°C, the inoculum was discarded, DMEM with 2% FBS was added, and the cells were maintained in a 5% CO2 environment. Cell culture flasks were examined daily for evidence of viral cytopathic effect (CPE). If no CPE was observed by day 7, the cultures were frozen and thawed and 500 μL of the supernatant was inoculated into fresh cell monolayers. If CPE was not observed after the third blind passage, samples were considered negative.
Results
Performance of real-time RT-PCR and RT-PCR protocols
All three lineage 1 and 2 isolates were detected successfully. The analytical sensitivity of the real-time RT-PCR was determined using 10-fold serial dilutions of RNA, extracted from the three isolates. The detection range of the assay was determined to be 1 TCID50/mL with amplification efficiency 95%. The two-step RT-PCR designed for the detection of WNV lineage 2 isolates proved to be specific for lineage 2 strains because it did not detect lineage 1 isolates.
Sentinel chickens
Eleven chickens out of 47 seroconverted to WNV, with ten seroconversions occurring between June and October in the western rural region of Thessaloniki and one seroconversion in September in the eastern region of the city. The first seroconversion occurred on June 29, 2011, in Delta municipality, in the town of Chalastra. To the west of Thessaloniki, the seroconversion rate remained stable with one new seropositive bird/week from July 13, week (W) 28, up to August 1 (W30), 2011 (Fig. 2), and peaked on August 17, 2011 (W33), with four new seropositive birds/week. Immediately after peaking, the seroconversion rate dropped to zero. The only seroconversion on the eastern suburban outskirts of the city occurred on September 21 (W38). The municipality with the highest rate of sentinel chicken seroconversion was Chalkidona (45%, 5 out of 11 chickens), followed by Delta (29%, 5 out of 17) and Thermi (6%, 1 out of 19) (Fig. 3). Serum neutralizing antibody titers to WNV ranged between 1:20 and 1:40 in all seropositive birds.

Weekly mosquito activity of the three most common Culex mosquito species on the western rural region of Thessaloniki (
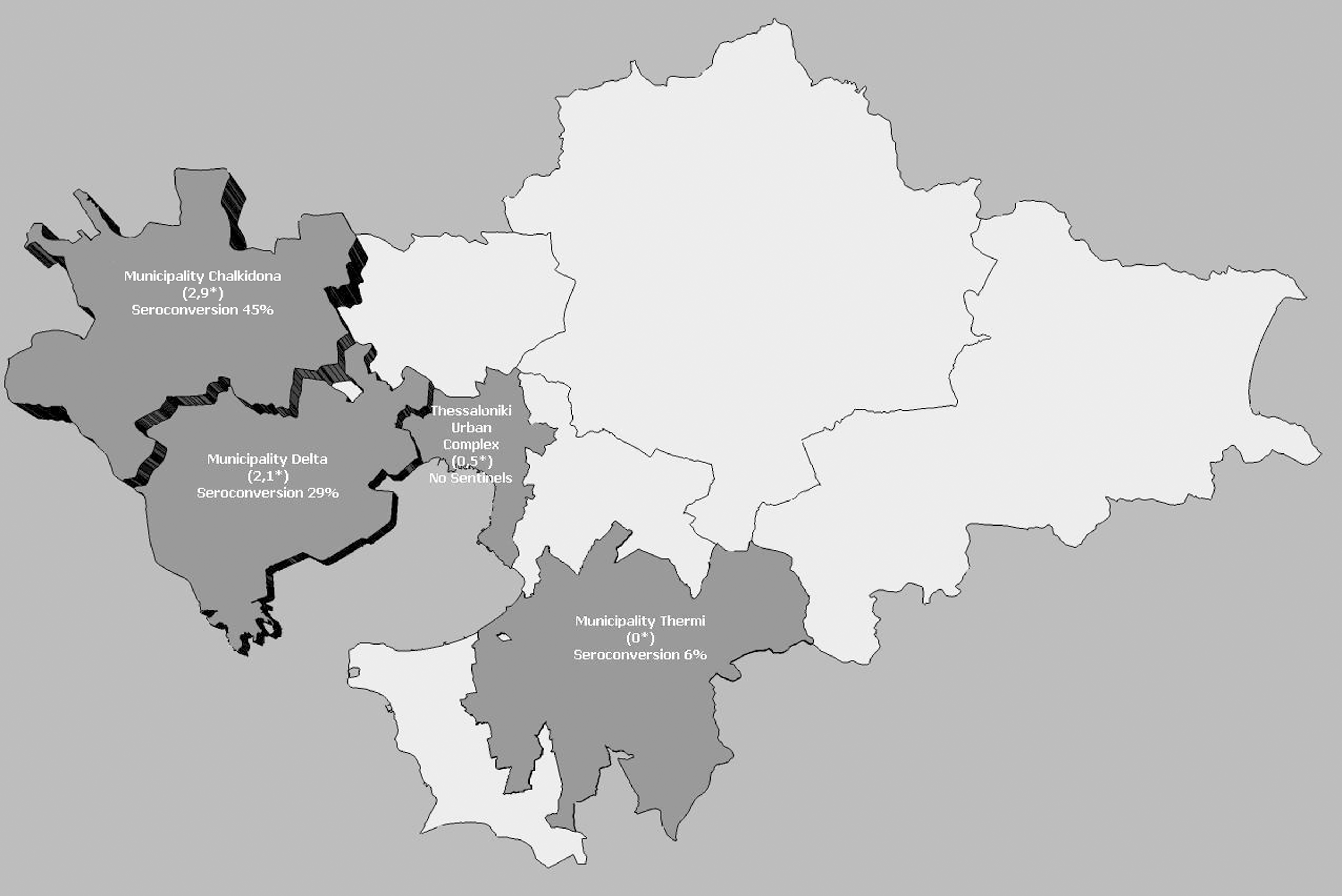
Chicken seroconversion percentage and human incidence rate [per 100,000 individuals (*)] for the municipalities of Chalkidona, Delta, Thermi and the urban complex of Thessaloniki. Incidence of human cases was provided by Greek CDC (personal communication, Marios Detsis, April 2012).
Retrospective examination of the plasma samples collected previously from the 11 seropositive chickens, using real-time RT-PCR, resulted in detection of WNV in two of them. Specifically, antibodies were detected in blood samples obtained on July 13 (W28) and August 17 (W33), whereas WNV was detected in samples taken 1 and 2 weeks before the detection of seroconversion in these birds, respectively. The full-length NS3 sequences of these two WNV isolates from chicken were determined to be identical (GenBank accession number JN398476) and presented the highest nucleotide sequence identity (99.9%) to the strain “Nea Santa-Greece-2010,” responsible for the 2010 Greek epidemic. The inferred NS3 amino acid sequences were 100% identical, maintaining the amino acid substitution H249P. Phylogenetic analysis revealed their high genetic relatedness to the less virulent lineage 2 strains of Hungary, 2004 (DQ116961) and Italy, 2011 (JN858070). However, all Greek isolates were clustered in a distinct phylogenetic clade, supported by high bootstrap values (Fig. 4).
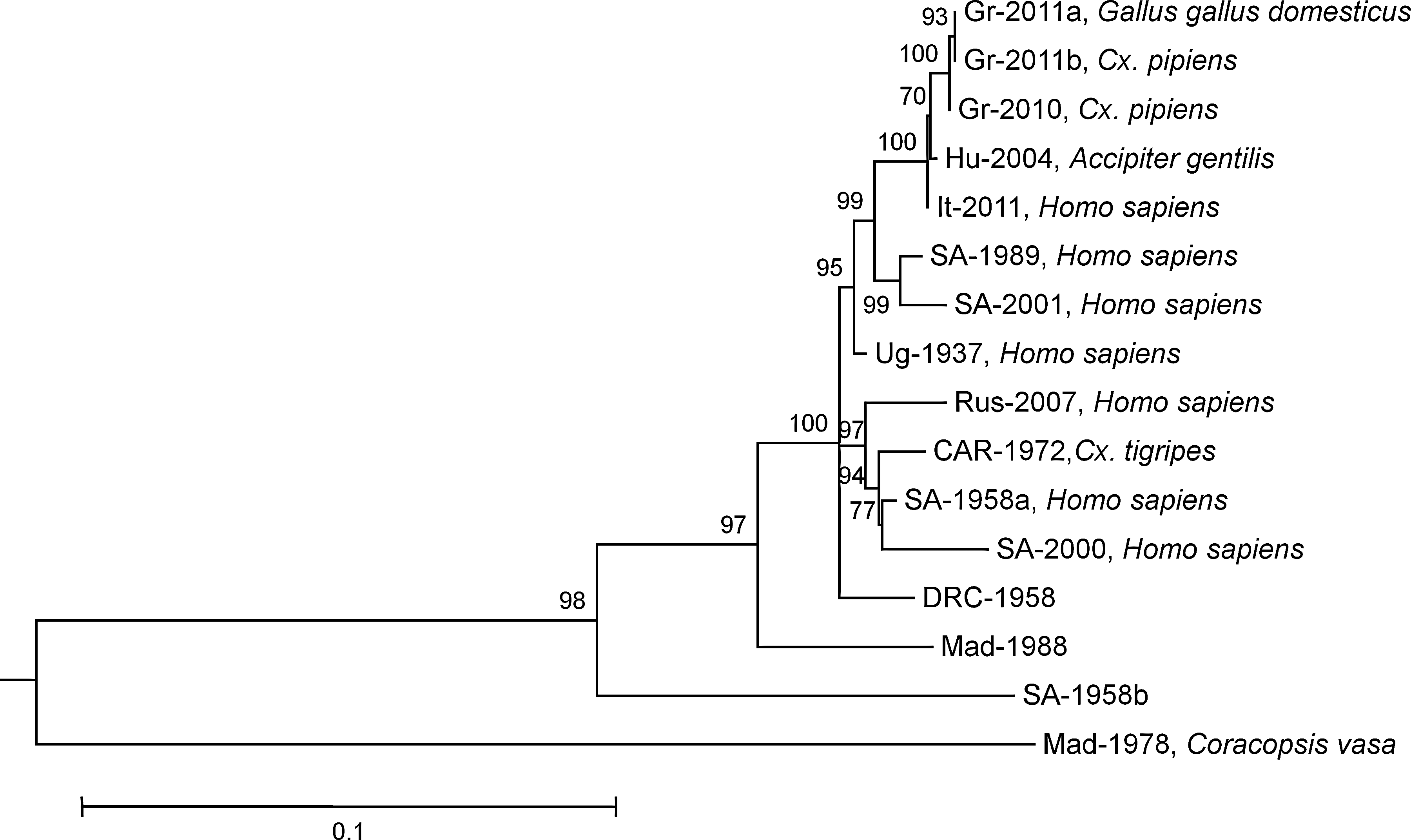
Phylogenetic tree inferred with maximum likelihood analysis, based on complete NS3 nucleotide sequences (1863 bp) of selected lineage 2 West Nile virus strains. Isolation source is indicated. Strain abbreviations (country, year, GenBank acc. no.)—Gr-2011a: Greece, 2011, JN398476; Gr-2011b: Greece, 2011, JX112013; Gr-2010: Greece, 2010, HQ537483; Hu-2004: Hungary, 2004, DQ116961; It-2011: Italy, 2011, JN858070; SA-1989: South Africa, 1989, EF429197; SA-2001: South Africa, 2001, EF429198; Ug-1937: Uganda, 1937, AY532665; Rus-2007: Russia, 2007, FJ425721; CAR-1972: Central African Republic, 1972, DQ318020; SA-1958a: South Africa, 1958, EF429200; SA-2000: South Africa, 2000, EF429199; DRC-1958: Democratic Republic of the Congo, 1958, HM147824; Mad-1988: Madagascar, 1988, HM147823; SA-1958b: South Africa, 1958, HM147822; Mad-1978: Madagascar, 1978, DQ176636. The tree was rooted using the "HNY1999" strain from human, acc. no. AF202541, which is a member of lineage 1, but has been removed to allow for better visualization of branch lengths. The general time reversible model with gamma distributed rates across sites and a fraction of sites assumed to be invariable (GTR+I+Γ), was selected as the best fitting nucleotide substitution model for the sequence data set. The numbers indicated on branches are nonparametric bootstrap (NPB) probabilities.
WNV successful amplification in Vero cells was confirmed by real-time RT-PCR. Final identification of the virus isolated was performed by sequencing of the NS3 gene.
Mosquito surveillance (infections and population dynamics)
Three Culex species (Cx. pipiens, Cx. modestus, Cx. theileri) were predominantly collected in CDC light traps in 2011. Cx. pipiens was ubiquitous, in contrast to Cx. modestus and Cx. theileri, which were collected only in the western and northeastern regions. In total 162,453 Culex mosquitoes were collected and, of those, 158,919 were collected from the western agricultural/rural region and the remainder from the northeast. Figure 2 presents the population dynamics of the most prevalent Culex species.
Cx. pipiens was the most abundant species overall and was trapped in residential areas, agricultural areas (rice fields), and natural wetlands. The population of Cx. pipiens started increasing early in the summer season on W23–W24. There was a distinctive peak on W26 in the western region, followed by a relatively stable period of activity with no major fluctuations until W35 when the population dropped and then gradually diminished. In the northeastern region, there were two notable peaks on W32 and W35 and on W38 a significant decrease on population activity was observed.
Cx. modestus was mostly associated with the rice fields as well as residential areas within the agricultural zone of west Thessaloniki. Their population started increasing significantly from W25–W28 and showed a distinctive peak on W29. After W29, the Cx. modestus populations started decreasing gradually and almost disappeared on W36. Cx. theileri was the least abundant mosquito and associated mostly with animal facilities in the eastern region. The species maintained a low and relatively stable activity throughout the sampling season.
A total of 150 pools of the three most prevalent Culex mosquito species were tested for WNV presence (Cx. pipiens 124 pools, Cx. modestus 23 pools, Cx. theileri 3 pools). One out of the 150 mosquito pools tested positive for the WNV “Nea Santa-Greece-2010” strain using the confirmation RT-PCR protocol. The NS3 sequence was identical to the one found in the chicken samples and was deposited into GenBank under accession number JX112013. Virus was detected in Cx. pipiens that were collected on July 23 (W25) from the traps located in the rice field region near the Axios Delta.
Dissemination of information to public health authorities to increase preparedness for vector control measures
On the first week of June (W22), Culex mosquitoes started to increase rapidly and authorities were notified. During the last week of June, Culex mosquitoes increased significantly, and the first chicken seroconversion occurred. Authorities were notified of each new seroconversion, their locations, and relevant Culex mosquito activities. Western rural areas were characterized by high mosquito activity and were considered as enzootic zones for WNV based on the numerous chicken seroconversions. Consequently, mosquito larviciding and public awareness measures were applied in those areas. Requests for registration of ultra-low-volume (ULV) aerial mosquito adulticiding products/techniques were submitted and registrations were approved on July 28. Aerial ULV adulticiding treatments were also conducted in Greece in 2010 to combat the WNV epidemic. In 2011, adulticiding was not initiated until the first human case was identified. Two aerial ULV treatments using pyrethroid insecticide (0.2% deltamethrin) were conducted over West Thessaloniki on W34 (Fig. 2) and covered ∼5000 hectares. Methodologies followed were described by Chaskopoulou et al. (2011b).
Discussion
Our monitoring program was successful at identifying enzootic transmission and allowing molecular characterization of WNV lineage 2 prior to the onset of human cases. A total of 10 confirmed human cases of WNV fever and neuroinvasive disease were reported by the Hellenic Center of Disease Control & Prevention (KEELPNO) in the Regional Unit of Thessaloniki. Six of those cases occurred within the urban complex of the city and the remaining four within the boundaries of Delta and Chalkidona municipalities in the western agricultural region of the city, where high enzootic WNV activity was observed as evidenced by the high number of chicken seroconversions. Incidence rates for WNND cases per 100,000 human population were 2.9 for Chalkidona, 2.1 for Delta, and 0.5 for the Thessaloniki urban complex. The chicken seroconversion rates paralleled the incidence of WNND cases for each municipality (Fig. 3). Further studies using more sampling points and birds are needed to confirm this possible correlation.
The first chicken seroconversion occurred ∼40 days prior to the first human case in the same rural region and ∼30 days prior to the first case within the urban region. Our results are in agreement with studies conducted in Florida, where the first WNV-positive human cases were detected 6.5 weeks after the first seropositive chicken (Blarkmore et al. 2003). On the other hand, in a recent study performed in Louisiana, human cases preceded sentinel chicken seroconversion (Unlu et al. 2009). Various factors are known to influence the capacity of sentinel chickens to function as early warning indicators. These include successful selection of virus amplification sites with high enzootic transmission, sufficient number of flocks and sentinels tested, as well as early detection of seroconversion by applying a sensitive diagnostic test.
In our case, we tried to further increase the analytical sensitivity of the commercial cELISA by lengthening incubation time after the addition of the sera and the conjugate, respectively. We observed that the absorbance signals increased significantly, and, most importantly, in a few of the infected birds tested, we managed to detect WNV-specific antibodies ∼1 week earlier, compared to the protocol without the modifications (data not shown). The specificity of the assay was not affected due to these modifications, since all the cELISA-positive results were confirmed by micro-PRNT and no false-positive results occurred. It is important to note that confirmation using a verification method, such as a serum neutralization test, should always follow the ELISA, because antigenic cross-reactions can occur with closely related flaviviruses.
WNV circulation has two basic types of cycles—a rural cycle (wild, usually wetland birds and ornithophilic mosquitoes) and an urban cycle (synanthropic birds and mosquitoes feeding on both birds and humans) (Hubalek and Halouzka 1999). During the Bucharest epidemic in 1996, the urban cycle predominated (Savage et al. 1996, Tsai et al. 1998). Nevertheless, our study and available observation from the Greek 2010–2011 epidemic suggested that the rural cycle was more important because people living in rural areas were twice as likely to develop WNND as persons living in urban areas (Danis et al. 2011). This is similar to studies in the United States and Canada, where WNND has a rural focus (Petersen and Hayes 2008).
Cx. pipiens and Cx. modestus were both highly abundant in the agricultural area of Thessaloniki, and both species are well-known vectors of WNV in Europe (Hubalek 2000). Out of the 150 Culex mosquito pools tested (including Cx. pipiens, Cx. modestus, and Cx. theileri) one positive pool of Cx. pipiens mosquitoes was collected near the Axios delta within the rice field region of western Thessaloniki. None of the mosquito pools collected near chickens that seroconverted tested positive for WNV. On the other hand, of the 32 Cx. pipiens pools tested from the rice field/river delta area one pool tested positive for WNV. The fact that we were able to detect infected mosquitoes with small sampling indicates that the rice field/delta region during the period of collection (July) had a very high concentration of infected mosquitoes. This wetland/agricultural area appears to serve both as WNV enzootic amplification foci as well as tangential transmission foci with the virus spreading to rural residential complexes and urban complexes as evidenced by: (1) High Culex mosquito populations observed in the rice field region west of Thessaloniki, (2) the infected Culex pool from the same area, (3) the high density of passerine and native birds associated with the delta in combination with the recent collection of a WNV infected Eurasian magpie from the region of Thessaloniki (Valiakos et al. 2011), and (4) the high enzootic transmission observed in sentinels, which was followed by human cases in the same residencies.
A drop in the adult mosquito population and decrease of WNV seroconversion rates in sentinel chickens followed emergency aerial adulticiding operations. No apparent changes in the temperature could account for such an abrupt drop. Furthermore, the mosquito populations on the east side of the city, where no spray was conducted, did not follow the same trends but rather showed an increase during the posttreatment season that resulted in the first positive chicken on mid-September. The efficacy of aerial ULV adulticiding in decreasing the number of infected WNV vectors has been demonstrated in several studies (Carney et al. 2008, Elnaiem et al. 2008, Lothrop et al. 2008, Macedo et al. 2010). However, the timing of vector control interventions can play an important role in reducing arboviral encephalitis transmissions. Emergency measures are largely ineffective if delayed until the index case appears (Connelly and Carlson 2009). If the aerial treatments had been conducted when the first Culex peak and chicken seroconversions were observed, then the incidence of human cases may have been reduced. Research on the timing of vector control measures should be implemented to refine our understanding of the most effective vector intervention strategies.
In our study, weekly sampling of captive chickens was successful for fast identification and isolation of the circulating WNV strain responsible for the 2010–2011 epidemic in Greece. However, we detected WNV in only 2 out of the 11 seroconverted chickens, probably due to the low duration of viremia (Langevin et al. 2011). Accordingly, we can conclude that if a similar surveillance system is targeting the isolation and molecular characterization of WNV, blood samples should be drawn at intervals shorter than 7 days. Epidemiologic observations indicate that the goshawk-Hungary-2004 strain circulated also in Austria during 2008 (Wodak et al. 2011) and has most likely spread south to Greece and Italy (Papa et al. 2011, Savini et al. 2012). However, only in Greece was the virus found responsible for the occurrence of a severe epidemic with high number of neuroinvasive disease cases in humans. The phylogenetic analysis presented here shows that all Greek isolates belong to a distinct recent evolutionary clade. The above data support the hypothesis that the “Nea Santa-Greece-2010” strain is a descendant of the goshawk-Hungary-2004 strain, which displayed higher virulence due to recent genetic changes. This hypothesis is further supported by a recent study, where evidence of local WNV enzootic circulation in wild birds prior to the 2010 human outbreak, as well as evidence of exposure of migratory wild birds to WNV prior to their arrival in resting areas of Greece during their autumn migration from central Europe is provided (Valiakos et al. 2012).
The surveillance system deployed demonstrated that the WNV lineage 2 strain responsible for the Greek 2010 was actively circulating in 2011 and probably managed to overwinter. The system proved valuable as an early warning indicator by detecting WNV enzootic activity 1 month prior to human cases and providing information regarding the circulating virus strain. As a result, public health authorities were informed promptly and control interventions were implemented. It also became clear that the rural agricultural region of Thessaloniki represents an important WNV transmission focus, providing additional evidence of a rural focus of WNV transmission in Europe. In conclusion, in Europe where numerous strains of different virulence coexist, molecular identification of the circulating viruses is a necessity and captive sentinel chicken surveillance with frequent sampling may be a very informative tool in accurately determining public health risk.
Footnotes
Acknowledgments
This project is funded by the Mosquito Control Program Contract of Thessaloniki County. The Development Agency of Thessaloniki (ANETH) is the implementation body of the program and the funding is provided by the Greek government, the Regional Authority of Central Macedonia, the Local Union of Communities and Municipalities of Thessaloniki County (TEDK) and the eight municipalities of Thessaloniki County. We thank Daniel Strickman and Orestis Papadopoulos for critical review of the manuscript, and Jonathan Day, Roxanne Connelly, and Gregg Ross for valuable discussions. Sylvie Lecolinet (European Reference Laboratory for Equine Diseases, UMR 1161 Virology, INRA-ANSES-ENVA, Maisons-Alfort, France) and Olga Mangana (Department of Virology, Athens Centre of Veterinary Institutes, Greek Ministry of Agriculture) are gratefully acknowledged for providing the PaAn001/France and Eg101/Egypt isolates. We thank George Ziankas for preparing the maps of Thessaloniki Region. We also thank all the residents of the Thessaloniki region who kindly accepted to host the sentinel chickens for the duration of the study. We thank the Deployed War Fighter Protection (DWFP) research program for supporting research at the USDA-ARS EBCL. Last but not least, we thank the Hellenic Center for Disease Control and Prevention for providing us with useful information.
Author Disclosure Statement
No competing financial interests exist.