
Abstract
The fat-tailed gerbil Pachyuromys duprasi is a common burrowing rodent found across the northern Sahara Desert from Morocco to Egypt. There is overlap in the geographical distribution and ecological habitats of P. duprasi, several Old World Leishmania species, and numerous sand fly vectors of Leishmania, but there are no records that document the natural occurrence of this gerbil with any species of Leishmania or phlebotomine sand fly. Experiments were conducted to determine its potential as a natural host and laboratory animal model for Leishmania major. Captive-born P. duprasi were inoculated subcutaneously (s.c.) in the tail with promastigotes or amastigotes of an Egyptian strain of L. major and monitored for signs of infection. Local swelling and erythema was visible 10–12 days after amastigote inoculation, and within 3–4 weeks swelling had increased tail widths by up to 78%. Infections progressed more slowly and less conspicuously following inoculation with promastigotes. Tissue density of amastigotes in the gerbil's tail lesions after inoculating with either stage of L. major was significantly lower than that produced in the footpads of BALB/c mice by the same parasite and incubation period. Laboratory transmission of L. major to P. duprasi by sand fly bite was demonstrated and acquisition of L. major, by bite, from tail lesions of infected P. duprasi to laboratory-reared Phlebotomus papatasi was also achieved with 10% of biting flies developing promastigote infections. The acquisition and development of L. major infections in P. papatasi after biting an infected P. duprasi and the susceptibility of P. duprasi to L. major delivered at low densities by sand fly bites indicate that fat-tailed gerbils could serve as a natural host and reservoir of L. major.
Introduction
The Syrian hamster and BALB/c mice are the most commonly used animal models for studying L. major, but there is no ideal laboratory animal for the xenodiagnosis and study of L. tropica, and there is uncertainty as to the variety and importance of different animal reservoirs of this parasite in nature (Jacobson 2004, World Health Organization 2007). Vector incriminations and quantifications of Leishmania infection risk are dependent upon the detection, isolation, and identification of Leishmania infections in human-biting, wild-caught sand flies, but even with well-advanced molecular-based identification systems, definitive, clinical, laboratory-based, and epidemiological studies are still dependent upon isolation and stable growth of the parasites in culture. Direct inoculations of parasites from individual sand fly infections into culture media often fail because of contamination, but direct in vivo inoculation of the parasites from dissected sand flies into a permissive laboratory animal yields an excellent success rate of isolation. This in vivo propagation and xenodiagnosis of Leishmania requires inoculation of footpads, nose, eyes, or tail base that causes stress and changes in the animal's behavior. An ideal permissive host would be docile and physically structured to promote both sand fly feeding and Leishmania lesion development over a wider, but less sensitive and non-load-bearing part of the animal's body (Felt et al. 2012).
The fat-tailed gerbil Pachyuromys duprasi Lataste, 1880 is a burrowing desert rodent found in the Western Desert of Egypt and across North Africa to Morocco (Osborn and Helmy 1980). The occurrence of geographically distinct P. duprasi subspecies in Egypt (P. d. natronensis), Libya (P. d. duprasi), and Algeria (P. d. faroulti), based on color shades of the animal's fur, is reportedly debated among taxonomists (Felt et al. 2012). Habitats of P. duprasi in the northern Sahara are the arid, sparsely vegetated sand sheets and rocky plains that occur inland from the more populated and agriculturally productive Mediterranean littoral zone. This gerbil is very docile, accepts a wide variety of foods in captivity, and has a high reproductive capacity (Felt et al. 2012). The conservation status of this animal in nature is listed as “least concern” (International Union for the Conservation of Nature 2012). The unusual, highly vascular tail of P. duprasi presents a larger and more sparsely haired surface than other rodent species and functions in the storage of water and fat. From the standpoint of blood feeding sand flies of the genus Phlebotomus, which are often found in close association with burrowing rodents, P. duprasi would appear to be an ideal host, providing flies with a large, easily accessed skin surface for blood feeding and a subterranean microenvironment for shelter and larval development. Although there is overlap in the geographical distribution and ecological habitats of P. duprasi, several Old World Leishmania species, and numerous North African sand fly species, there are no records of fat-tailed gerbils serving as a host or natural reservoir of any Leishmania species, and there are no published accounts of sand flies naturally associated with this common mammal. We considered P. duprasi to possess ideal qualities for laboratory study of Leishmania transmission by sand flies and devised simple trials to evaluate this animal's potential as a laboratory model and natural host for L. major. Initial success with studies of L. major in P. duprasi, would provide impetus and guidance toward an assessment of L. tropica and other Leishmania species in this animal. The objectives of this initial study were to: (1) Determine the time course and manifestation of L. major infection in P. duprasi by subcutaneous (s.c.) inoculation of either amastigote or promastigote stages of the parasite and (2) evaluate this animal as a laboratory model for L. major transmission studies with the blood-feeding sand fly P. papatasi.
Materials and Methods
Laboratory animals
The Egyptian subspecies P. d. natronensis, de Winton, 1903 used in this study was originally collected from the Western Desert of Egypt in 1996 and established from 24 wild-caught “founders” as a laboratory breeding colony at Naval Medical Research Unit #3 in Cairo, Egypt (Felt et al. 2008, Felt et al. 2012). All animals used in this study were born and raised in captivity under humane conditions and in an Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC)-accredited facility. Male (n=14) and female (n=14) animals, 8–10 weeks old weighing 70–90 grams, were used. BALB/c mice 4–6 weeks old and of either sex served as positive controls/comparator hosts. The research plan and procedures were approved by a duly constituted Institutional Animal Care & Use Committee (IACUC). Despite a subspecies pedigree of P. d. natronensis, this report will refer to them generally as “gerbils,” “fat-tailed gerbils,” or P. duprasi.
Parasite strain
The parasite strain, L. major (IPAP/EG/1989/SI-177) was isolated from wild-caught female P. papatasi from the Northern Sinai, Egypt, and held in liquid nitrogen (Mansour et al. 1991). Promastigotes were thawed and cultured in Schneider's Drosophila medium to stationary phase for inoculation of gerbils and mice. Amastigote stage L. major parasites for inoculation were produced in BALB/c mice.
Infection by inoculation of promastigote- or amatigote-stage parasites
Aliquots of cultured metacyclic-stage promastigotes (0.25 mL containing 2.1×106 parasites) were injected s.c. into the dorsal tail surface of P. duprasi (n=14), and a reduced volume of 0.025 mL containing 2.1×105 promastigotes was inoculated into the smaller left hind footpads of BALB/c mice (n=4). Alternatively, amastigote-infected tissue excised from a donor animal was homogenized in sterile phosphate-buffered saline (PBS) and counted in a hemocytometer; 0.25 mL of this suspension containing 2.6×106 amastigotes was inoculated into the dorsal tails of P. duprasi (n=14). A volume of 0.025 mL (2.6×105 amastigotes) was inoculated into the left hind footpads of BALB/c mice (n=3). Animals were monitored daily for signs of infection at the sites of inoculation. Gerbil's tail widths were measured once weekly with vernier calipers and recorded. Animals were euthanized 30–47 days after infection and necropsied. Spleen, liver, bone marrow, and lymph nodes from gerbils were cultured individually in NNN medium and checked for growth of L. major parasites over 3 weeks. Tissue samples from uninfected controls, amastigote-infected gerbils (tails), and amastigote-infected BALB/c mice (footpads) were fixed in formalin, stained with Hematoxylin & Eosin (H&E), and examined by light microscopy. Descriptive statistics (means+95% confidence intervals) and t-tests were used to determine the significance of differences in P. duprasi tail widths, time to maximum swelling of the infected tails, and percent change from baseline in tail widths.
Quantifying amastigote tissue infections
Subsamples of gerbils (n=7) and BALB/c mice (n=7) were euthanized after lesions developed from amastigote (day 21) and promatigote (day 47) inoculations. Amastigotes were isolated from the infected tail and footpad tissue according to Bastien and Killick-Kendrick (1992). From these isolates, amastigote counts for each animal were determined by light microscopy and expressed as counts/mL according to the published polystyrene microbead method of Cenini et al. (1989).
Infection and transmission studies with blood-feeding sand flies
A laboratory colony of P. papatasi that originated in North Sinai, Egypt, was used for infection and transmission studies. Colonized sand flies were reared and maintained at 27°C, 90% relative humidity, under a 12:12-h light:dark photoperiod, and adult flies were provided 30% sucrose solution as a carbohydrate food source except when females were fasted prior to blood feeding. Adult female flies 5–6 days old were fasted for 24 h before blood feeding for 1 h on the tails of infected P. duprasi. Flies that fed were dissected at time points postfeeding to determine the acquisition and development of L. major. To demonstrate the transmission of L. major by bites from sand flies to naïve P. duprasi, laboratory-reared 5- to 6-day-old female P. papatasi were initially infected by membrane-feeding on a suspension of amastigotes in hamster blood and permitted to refeed 14 days later on a naïve P. duprasi, selecting their feeding sites freely on the anesthetized animal. Flies that refed were dissected to verify and quantify their infectivity. The gerbil was monitored weekly for signs of Leishmania infection. Suspected cutaneous lesions were sampled by aspiration biopsy, and Giemsa-stained preparations were examined by high-power light microscopy. This animal was euthanized 6 weeks after infective sand fly biting and necropsied. Samples of spleen, liver, bone marrow, and lymph nodes were cultured individually in NNN medium and checked for growth of L. major parasites over 3 weeks.
Results
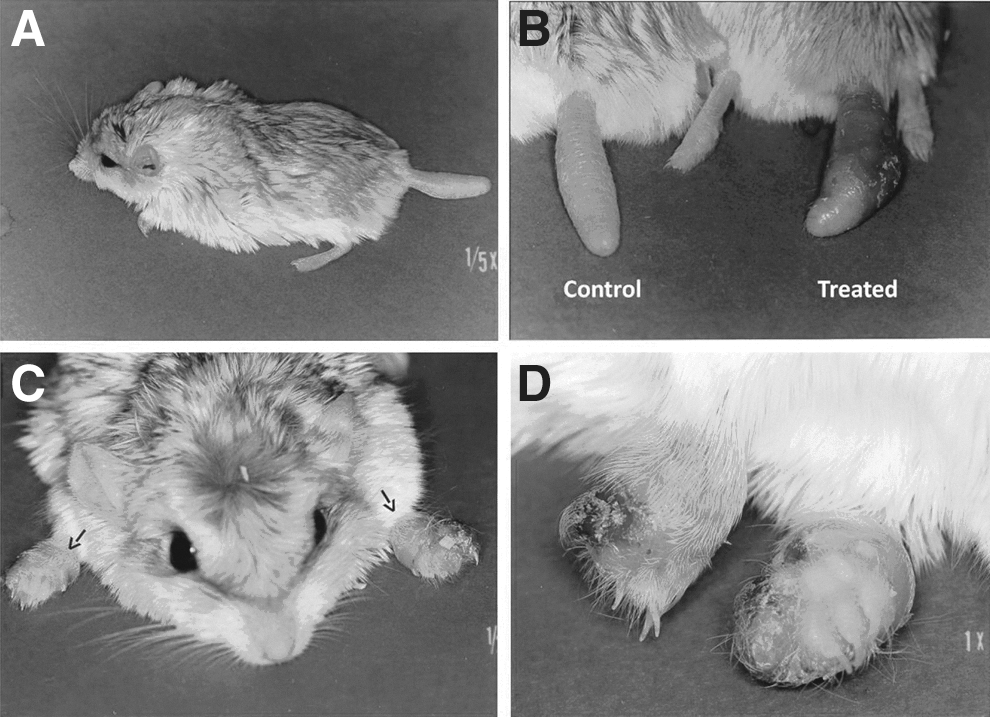
Following s.c. inoculation of L. major amastigotes into their tails, all gerbils developed signs of infection, first noted on days 10–12 as swelling at the sites of injection. Dermatological manifestations of infection changed over time, beginning as erythema and redness at the site of inoculation and progressing to superficial dermatitis and to an ulcerative lesion by days 18–24 (Fig. 1B). Preulcerative swelling was variable among gerbils, ranging from as little as 10% (0.7 mm) to 78% (4.3 mm) over baseline, and with a mean day-21 increase over baseline tail width of 35.5% (2.4 mm) (Table 1a). BALB/c mice that received a 10-fold lower dose of the amastigotes into their dorsal footpads demonstrated similar progression, with first signs of swelling on days 9–13, followed by dermatological lesions and ulceration by days 20–29.
(
s.c., subcutaneous; CI, confidence interval.
Subcutaneous inoculation of metacyclic-stage cultured promastigotes also led to infection and swelling in all gerbils, with wide variability among individuals (Table 1b). The day-21 mean increase over baseline tail width following promastigote inoculation was 21.1%, ranging from 2.5% to 49%. Table 1 shows that the mean day-21 response to amastigote inoculations was significantly greater than that induced by promastigotes (mean increase, 2.4 mm vs. 1.5 mm; p=0.04; mean % increase, 35.5% vs. 21.1%; p=0,03). There was no further swelling beyond day 39 postinfection in 5 of 10 gerbils that received promastigotes, but one other animal demonstrated a rapid 29% increase in swelling between days 26–32. Table 2 shows that mean time in days required for amastigote inoculations to induce maximum swelling of gerbils' tails was significantly reduced from that of promastigotes (21.5A days vs. 34.4P days; p<0.001). The mean maximum tail widths and percent increase attained was comparable in the two groups. Figure 1B shows the significant swelling and ulceration that was apparent in the tail of a gerbil that was injected s.c. with amastigotes 21 days earlier. Lesions did not appear at any other location, and in 7 of 11 animals no further swelling of the tail was detected beyond day 19 postinfection.
s.c., subcutaneous; CI, confidence interval.
Examination by microscopy of H&E-stained lesion tissue taken on day 21 from representative animals showed moderately, diffuse, granulomatous dermatitis, with intrahistiocytic protozoal organisms, etiology consistent with Leishmania infection, in the gerbils and mice. There were abundant infected macrophages filling the connective tissue spaces of gerbils and mice, but connective tissue was considerably reduced in gerbils, owing to the preponderance of adipocytes. Accordingly, counts of L. major amastigotes isolated from footpad lesions were greater than those from tail lesions in six of seven comparisons. In this small paired sample (Table 3), collectively for both promastigote- and amastigote-induced lesions, the mean number of amastigotes isolated from mouse footpad tissue was significantly greater than that isolated from gerbil tail tissue (1.2×107/mL vs. 4.0×106/mL; p=0.003). Transmission of L. major to P. duprasi by bite of infective P. papatasi was demonstrated. Despite no prior exposure to P. duprasi as a blood source, colony-reared, female P. papatasi that were earlier infected by membrane feeding (101/146=69% initial feeding rate) refed readily on a naïve gerbil (20/30=67% refeeding rate). Subsequent dissection of the 20 refed flies determined that 19 (95%) were infected with promastigotes and probably infective. Twenty-three days after being bitten by these L. major–infective sand flies, cutaneous ulcers developed on both of the gerbil's forefeet (Fig. 1C, D). The animal was monitored for another week, but no other lesions developed. Microscopy of the Giemsa-stained lesion aspirates confirmed transmission of parasites by sand fly bite and localized progression of the infection to the tissue amastigote stage with typical manifestations of CL pathology. Cultures of necropsied organs from this infected gerbil grew no Leishmania parasites.
Gerbils: 2.1×106 promastigotes/animal or 2.5×106 amastigotes/animal.
BALB/c mice: 2.1×105 promastigotes/animal or 2.5×105 amastigotes/animal.
s.c., subcutaneous; CI, confidence interval.
Transmission of L. major from an infected gerbil to uninfected blood-feeding sand flies was also successfully demonstrated. When restricted to feeding only upon the gerbil's infected tail, 65 of 119 (54.6%) female sand flies engorged within a 1-h feeding period. However, a relatively low infection rate of 10% (2/20) was determined for sand flies that fed on the gerbil's swollen tail. The two positive flies were dissected on days 5 and 8 postfeeding and contained moderate promastigote infections (>100 parasites) in their abdominal and thoracic midguts.
Discussion
The Egyptian fat-tailed gerbil P. d. natronensis demonstrated susceptibility to infection by L. major at the low levels delivered by sand fly bite. Inoculation by syringe of gerbil's tails with either promastigotes or amastigotes developed into a localized cutaneous lesion within a time span comparable to that of BALB/c mice and attained sufficient parasite density to infect biting sand flies. Collectively these findings indicate that fat-tailed gerbils could serve as natural hosts of L. major and may be a useful new animal model for studies of Leishmania infection and pathogenesis.
Although golden hamsters and BALB/c mice are the classic susceptible animal models employed for studies of L. major infection and pathogenesis, the P. duprasi model allowed us to monitor and quantify progression of the infection from the day of inoculation by simple caliper measurements of tail width. Infection progressed in all animals to an open cutaneous ulcer at the injection site 3–4 weeks after amastigote inoculation or 4–6 weeks after promastigote inoculation. Considerable variation was observed among inoculated gerbils in the appearance, duration, and extent of swelling and the ulcerative skin lesions. Such differences are typically attributed to infective dose, parasite strain differences, and genetic constitution of the host (Mauel and Behin 1987). Although L. major, infections would be expected to worsen progressively, metastasize, and ultimately kill BALB/c mice, we did not allow infections in P. duprasi (or mice) to progress beyond day 47 and consequently could not determine whether infections were self-healed or lethal in gerbils.
The apparent cessation of further swelling that was observed in more than half of the gerbils suggests an immunological capacity to control, or even resolve the infection and may be indicative of a well-developed evolutionary relationship and adjustment between P. duprasi and L. major. The greater yield of amastigotes from footpad lesions of BALB/c mice as compared to infected tissue from P. duprasi tails is considered to reflect the extreme susceptibility of the mouse strain and its inbred immunologic deficiencies, but may also relate to simple anatomical and physiological differences at the injection site. Histologically, the gerbil's tail, as compared to the mouse footpad, lacked apocrine glands, was less vascularized, and contained mainly compacted adipocytes with reduced interstitial connective tissue. The abundant adipose tissue of the gerbil's tail, comprising 70–75% of the tail in cross section, provides a critical survival function in wild P. duprasi as a caloric and nutritional reserve. Its anatomical bulk also contains water and serves as an insulator, maintaining warmer surface and internal temperatures for its tail than cooler extremities like ears, nose, and feet. These site-specific physico-chemical and nutritional conditions may be important limiting factors for the propagation of L. major. Nevertheless, P. duprasi tail tissue was clearly susceptible to rapid, extensive pathology following inoculation of L. major promastigotes, and the amastigote density achieved there was both sufficient and functionally capable of infecting blood-feeding sand flies.
The rapid swelling and ulceration of gerbil forepaws that resulted from sand fly bites that injected far fewer parasites than were injected by syringe into tails suggests that the forepaws and feet of P. duprasi are more ideal sites for L. major infection and propagation, but consideration should be given to the likelihood that this was an exaggerated host effect in response to massive, first-time exposure to potent immunosuppressive components in the sand fly saliva. If natural populations of fat-tailed gerbils coexist with, and are routinely exposed to sand fly biting, this pre-exposure would presumably arm them with significant tolerance and/or immunity to L. major infections (Belkaid et al. 1998).
Screening by microscopy and culture failed to produce any evidence of parasite dissemination to primary lymph nodes, spleen, liver, or bone marrow in either P. duprasi or the BALB/c controls. This may be due to a particular characteristic of the parasite strain used, or to the missing contribution of sand fly salivary factors, but we concede that detection of parasite DNA by PCR is necessary to rule out the distribution and persistence of L. major in P. duprasi. It is notable that intradermal inoculation of L. major promastigotes into the ears of two inbred murine strains (BALB/c and B10D2) led to self-limiting dermal lesions at the inoculated sites, but PCR determined that parasites were rapidly disseminated and were persistent for at least 12 months near the site of inoculation and its draining lymph node (Nicolas et al. 2000). In Syrian hamsters, s.c. inoculations of L. major promastigotes into the footpads led to disseminated cutaneous lesions in 38% of the animals with parasite DNA initially detected in blood after 1 month and subsequently in spleens and livers 3–10 months postinoculation (Soliman 2006).
The location, progress, and extent of lesion development by L. major in a mammalian host is governed by a complex interplay of vector, parasite, and host factors, but wild rodents that serve as reservoirs of L. major typically develop lesions and concentrations of the parasites on their nose, ears, and feet. All sites that present skin surfaces most free of hair and easily accessible to sand fly blood feeding (Killick-Kendrick and Ward 1981). These areas are also highly vascular, with abundant blood vessels. Despite their rich blood supply, these extremities, provide a cooler environment that may be conducive to the parasite, either by virtue of “dampened” host immune mechanisms (Scott 2007, Von Stebut 2007), or through innate temperature sensitivities of the parasite that underlie its tissue tropism (Callahan et al. 1996).
Although the tail of P. duprasi seemed to be a less ideal site for propagation of L. major, it was apparently also a less desirable location for sand fly blood-feeding. Fasted P. papatasi blood-fed avidly and successfully on P. duprasi when only the tail was accessible, but refeeding sand flies given free access to all parts of the gerbil showed no special attraction to the tail and fed preferentially on the feet, nose, and ears, the same sites they selected in BALB/c mice. It was not unusual that only two of 20 blood-fed sand flies acquired and developed L. major infections after feeding on the gerbil's tail, but it did seem unusual that only 20 of 65 (31%) first-time blood-fed flies survived the 7 additional days needed to mature parasites to the promastigote stage. However, this was a single observation and may be due to chance, or poor husbandry, rather than to a factor present or missing in the blood.
In summary, this study indicates that P. duprasi may be a suitable host for blood-feeding sand flies and does have the capacity to serve as a zoonotic host or reservoir of L. major. The unusual tail of this animal, while appearing to make it more vulnerable, seems not to serve as an ideal site for either sand fly feeding or propagation of the parasite. Furthermore, a molecular-based study is warranted to accurately determine whether P. duprasi can support broad dissemination and long-term, but relatively disease-free, maintenance of L. major and other Leishmania species.
Footnotes
Acknowledgments
The authors thank Dr. Stephen A. Felt (Laboratory Animal Medicine Residency Program, Stanford University, Stanford, California) for his critical reading and comments on the manuscript and also Maria Badra of U.S Naval Medical Research Unit No. 3, Vector Biology Research Program for reviewing and processing this manuscript.
Author Disclosure Statement
The views expressed in this article are those of the authors and do not necessarily reflect the official policy or position of the Department of the Navy, Department of Defense, nor the U.S. Government. One co-author is a military service member; the first author is an employee of the U.S. Government. This work was prepared as part of our official duties. Title 17 U.S.C. §105 provides that ‘Copyright protection under this title is not available for any work of the United States Government’. Title 17 U.S.C. §101 defines a U.S. Government work as a work prepared by a military service member or employee of the U.S. Government as part of that person's official duties.
The experiments reported herein were conducted in compliance with the Animal Welfare Act and in accordance with the principles set forth in the “Guide for the Care and Use of Laboratory Animals,” Institute of Laboratory Animal Resources, National Research Council, National Academy Press, 1996. The research protocol and methods employed in this work were reviewed and approved by a duly constituted Institutional Animal Care and Use Committee (IACUC), NAMRU-3 Animal Use Protocol No. 97-02. All animals used in this work were cared for and maintained under conditions approved by the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC). This work was supported by Work Unit Number: CRADA-6.1-U.