
Abstract
In a previous study, our group investigated the Babesia spp. prevalence in questing Ixodes ricinus ticks from nine city parks in South Germany in the years 2009 and 2010. We showed predominant prevalence of B. venatorum (in previous literature also known as Babesia sp. EU1), especially in those parks in a more natural condition and with occurrence of large wild animals, such as roe deer. To obtain longitudinal data and to broaden the knowledge about this pathogen, further investigations were carried out in 2011 and 2012 in four of those city parks. Two additional habitat types were chosen for comparison of prevalence data and species analysis focusing on occurrence of potential reservoir hosts. A total of 10,303 questing I. ricinus were collected in four city parks, a pasture, and a natural area in Bavaria, and a representative number of samples were investigated for prevalence of DNA of Babesia spp. (n=4381) and Rickettsia spp. (n=2186) by PCR. In the natural and pasture area, a significantly higher Babesia spp. prevalence compared to the urban area was detected. The natural area revealed sequences of B. microti, B. venatorum, and B. capreoli. In the pasture and urban habitat, predominantly B. venatorum was found, whereas B. capreoli was less frequent and only one B. microti–infected tick was found. All B. microti sequences were 100% identical to the zoonotic Jena/Germany strain. For Rickettsia spp., the significantly highest prevalence was also detected in the natural and pasture areas, whereas lower prevalence was found in the urban area. Sequence analysis revealed R. helvetica (98%) and R. monacensis (2%). Prevalence rates and occurrence of Babesia spp. and Rickettsia spp. differed in urban, pasture and natural sites, most likely depending on the habitat structure (natural or cultivated) and therefore on the appearance and availability of reservoir hosts like roe deer or small mammals.
Introduction
Rickettsia spp. cause tick- or flea-borne rickettsioses in humans (Parola et al. 2005). R. helvetica and R. monacensis were previously detected in Ixodes ricinus in urban and natural areas in southern Germany with prevalence ranging from 4% to 13% depending on stages (Silaghi et al. 2008, Schorn et al. 2011b). The vertebrate reservoir host for Rickettsia spp. remains unclear, but rodents and other small mammals have been proposed for R. helvetica and R. monacensis (Burri et al. 2011, Schex et al. 2011). Birds have also been discussed as possible reservoir hosts for Rickettsia spp. (Santos-Silva et al. 2006, Stańczak et al. 2009). The pathogenicity of R. helvetica in humans is still in dispute, but was mentioned in context with febrile illness, meningitis (Nilsson et al. 2010), and sudden cardiac failure (Nilsson et al. 1999, Fournier et al. 2000). R. monacensis was reported as causative agent of Mediterranean spotted fever–like illness in Italy (Madeddu et al. 2012).
The aims of this study were the comparison of the: (1) Occurrence of Babesia spp. in 2011 and 2012 in questing I. ricinus from city parks with previous results from 2009 and 2010 (Schorn et al. 2011b) and (2) prevalence and species diversity of Babesia spp. and Rickettsia spp. in questing I. ricinus ticks collected from areas with heterogeneous habitat structures and reservoir hosts
Materials and Methods
Study sites
Six study sites in South Germany were investigated in the years 2011 and 2012. The sites included four urban areas located in the cities of Munich (M2, M3) and Regensburg (R1) and close to Lake Starnberg south of Munich (B) as previously described (Schorn et al. 2011a, b), one pasture area between Lake Starnberg and Lake Ammersee (‘Kerschlach’, K), and one natural forest area about 44 km south of the city of Augsburg (‘Angelberger Forst’, T). Detailed descriptions of geographic data and a map showing the location of study sites have been published elsewhere (Overzier et al. 2013a). Because the present study pursues a different focus than the studies previously investigating city parks of the urban area (Schorn et al. 2011a, b), the study site description of the urban sites is extended.
Urban area
Sampling sites M2 (‘English Garden’, 375 ha) is a large city-center park divided by a road into a northern and a southern part. The southern part, where sampling collection took place, is under strong anthropogenic influence, highly frequented by visitors and dogs, maintained by gardening (e.g., mowing), and intensively used in summer with permission for park visitors to leave the walkways. Interaction between domestic and wild animals is limited to mice, hedgehogs (Erinaceus europaeus), foxes (Vulpes vulpes), birds, and so on, as big wild mammals such as roe deer (Capreolus capreolus) or wild boar (Sus scrofa) are nonexistent in this part of the park.
Sampling site M3 (‘Nymphenburger-Schlosspark’, 141 ha) is a nature-orientated city park, surrounded by park walls, and included in a nature conservation program. More forest-like structures with higher tree density, scrub, and wild animals such as roe deer exist in this park. Grass areas in M3 are extensively used and less frequently mowed, and therefore areas with longer grass exist and park visitors have to stay on the walkways.
Sampling site R1 (‘Dörnbergpark’, 7.4 ha) is located in the city center of Regensburg and surrounded by a wall. Visitors are allowed to leave the walkways and to use the mowed grass areas in summer. Due to the small size and the inner city location of sampling site R1, there is no occurrence of big mammals like roe deer or wild boar. Twenty-six different bird species are common in this park and for 20 bird species it is an important habitat for brood (Stadt Regensburg Umweltreferat-Gartenamt 1996).
Sampling site B (‘Schlosspark-Berg’; 30 ha total, of which ∼18 ha is public and ∼12 ha private ) is a forest-like park on the eastern shore of Lake Starnberg with scrub and walkways. Large wild mammals such as roe deer and foxes exist and hunting is practiced. Dogs need to be kept on the leash in sampling sites M3, B, and R. Deciduous trees and bushes exist in all investigated city parks (Schorn et al. 2011a, b).
Pasture area (site K)
Two fenced pastures (each ∼8 ha) used for stock breeding of cattle surrounded by a forest area were investigated. Questing ticks were collected inside the fenced area next to the forest. Contact between wild animals and domestic cattle exists because animals from the forest can enter the pasture.
Natural area (site T)
The ‘Angelberger Forst’ (641 ha), a forest that is also included in a nature conservation program with occurrence of several wild animals, with very low frequency of visitors and therefore little to no interaction between wild and domestic animals, was investigated. This area has been described in detail before (Overzier et al., 2013b).
Collection of samples
In April, May, and June of 2011 and 2012, respectively, questing ticks were collected with a 1-m2 white cotton flag once a month at each study site and stored in 70% ethanol. For measuring the tick abundance, an area of 300 m2 was flagged, sectioned in three transects of 100 m2 each. The objective was to collect a minimum of 40 ticks of each gender and stage (without larvae) per site and month. If the number of collected ticks on 300 m2 was too low, additional ticks were collected nearby each transect to reach the mentioned number. The ticks were washed twice with distilled water, identified (Hillyard 1996), and stored at −20°C until further investigation as described (Schorn et al. 2011a). Larvae were included in this study in pools of up to five ticks because transovarial transmission is known for large Babesia spp. (Homer et al. 2000, Bonnet et al. 2007a, b) and Rickettsia spp. (Socolovschi et al. 2009). Results of the tick density per site and month and relationship of stages have been published elsewhere (Overzier et al. 2013a).
DNA extraction, PCR amplification, and sequencing
DNA extraction was performed with the automated Maxwell® 16 System (Promega, Mannheim, Germany) from a maximum of 60 questing ticks of each developmental stage, gender, and month. DNA concentration was measured with a spectrophotometer (Nano Drop® ND-1000, PeqLab Erlangen, Germany). All samples with a DNA concentration ≥10 ng/μL were diluted 1:4 with elution buffer to raise the volume. This step was done to avoid false-negative results caused by highly concentrated DNA inhibiting the PCR process. DNA concentration used for PCR reached from 1.8 ng/μL up to 61.4 ng/μL with a mean concentration of 12.0 ng/μL. Details of the DNA extraction process have been published elsewhere (Overzier et al., 2013b).
The 18S rRNA gene of Babesia spp. and the gltA and ompB genes of Rickettsia spp. were detected by conventional PCR, and species identification was carried out by sequencing and sequence analysis as previously described (Schorn et al. 2011b). The PCR for Babesia spp. DNA was carried out in ticks from 2011 and 2012, whereas PCR for Rickettsia spp. was only performed in ticks from 2011.
Statistical analysis
Two logistic regression models were estimated to investigate the effect of gender, month, and year on the probability of positivity. One model also included habitat, the other one the different sites. A simultaneous test for general linear hypotheses based on multiple comparisons of means with Tukey contrasts (Hothorn et al. 2008) was used to test for differences between gender, month, habitat, and site, respectively. Statistical analysis was performed accepting the minimum prevalence (in a positive pool of larvae, one tick was assumed as positive). The variable “sampling site R1” was removed for the statistical analysis of Babesia spp. because no infection was seen on this sampling site and therefore the estimation of the parameters was not possible. Statistical analysis was performed with R version 2.15.1 (R Development Core Team, 2012).
Results
In all, 2186 I. ricinus ticks in 2011 and 2195 in 2012 were investigated.
Prevalence of Babesia spp. in I. ricinus
A total of 1.0% (45/4381) of investigated questing ticks was positive for DNA of Babesia spp. in 2011 and 2012. The following prevalences were detected in the different habitat types (Table 1): (1) Urban area, adults, 0.3% (3/889), nymphs, 0.0% (0/500) in 2011 and adults, 0.5% (5/947), nymphs, 1.5% (7/464) in 2012; (2) pasture area, adults, 1.8% (4/225), nymphs, 2.1% (3/140) in 2011 and adults, 1.3% (3/234), nymphs, 0.8% (1/120) in 2012; (3) natural area, adults, 2.5% (2/79), nymphs, 3.3% (4/120) in 2011 and adults, 3.1% (7/226), nymphs, 3.3% (4/120), larvae, 2.5–12.5% [(2–10/80) including two positive pools] in 2012.
A, adults; N, nymphs; L, larvae.
Urban, 11x B. venatorum, 8x M3, 3x B, 1x B. microti strain Jena/Germany (M2), 3x B. capreoli (2x M3,1x B); pasture, 9x B. venatorum, 2x B. capreoli; natural, 7x B. venatorum (including 2 pools of larvae), 11x B. microti strain Jena/Germany, 1x B. capreoli.
47x R. helvetica (29x urban, 9x pasture, 9x natural), 1x R. monacensis (study site B).
n, number of larvae pooled.
nd, not done.
There was no statistically significant difference in prevalence between stage and gender, as well as between months (data not shown) and years (Table 1). The natural (p<0.001) and the pasture habitat (p<0.05) showed statistically significantly higher prevalence than the urban habitat. The prevalence on study site ‘T’ (natural) was significantly higher than on study sites ‘M2’ and ‘B’ (urban) (p<0.05).
18S rRNA gene
All Babesia spp. positive samples were sequenced (Fig. 1). Overall 27x B. venatorum, 6x B. capreoli, and 12x B. microti strain Jena/Germany were detected by comparison with GenBank sequences. Distribution in the different habitats was as follows: (1) Urban area, 11x B. venatorum (sites: 8x M3, 3x B), 3x B. capreoli (sites: 2x M3, 1x B), 1x B. microti strain Jena/Germany (site M2); (2) pasture area, 9x B. venatorum, 2x B. capreoli; and (3) natural area, 7x B. venatorum (including 2 pools of larvae), 1x B. capreoli, 11x B. microti strain Jena/Germany.
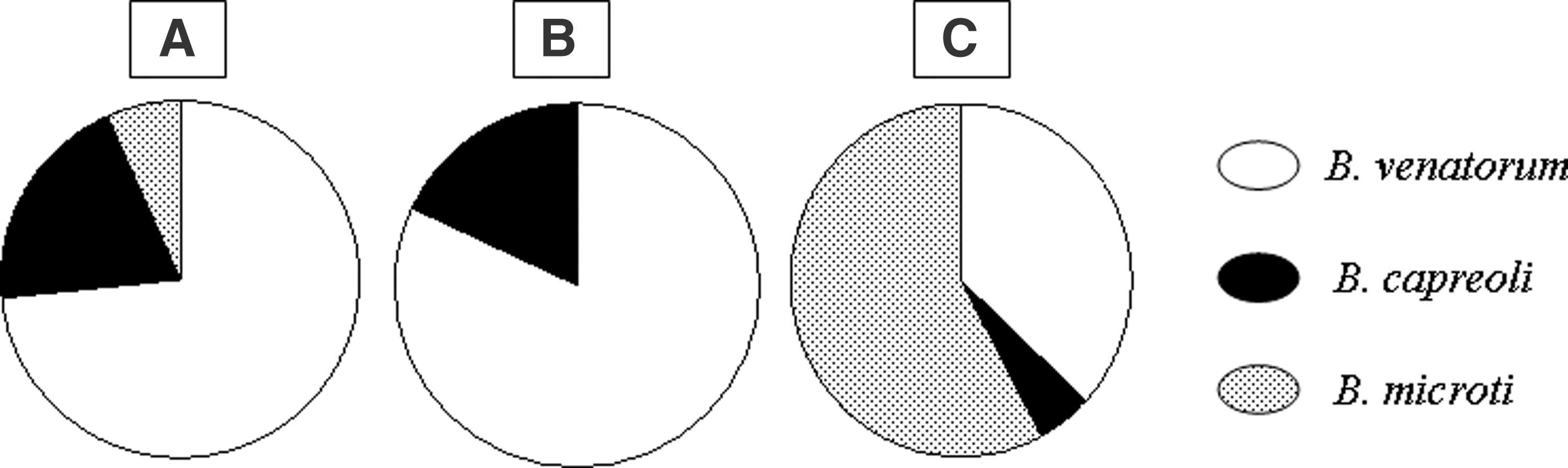
Prevalence of Babesia venatorum, B. microti, and B. capreoli. (
B. venatorum sequences were 99–100% identical to sequences previously found in roe deer in the natural area (GenBank accession nos. JX627351, JX627352) and in I. ricinus ticks from the previous study in the city parks (GenBank accession nos. JN036420–JN036426).
B. capreoli sequences showed 100% identity to sequences from I. ricinus and roe deer previously found in the natural area (GenBank accession nos. JX627350, JX627353, JX627354, JX627358). One sequence found on the pasture was 99% identical to both B. capreoli and B. divergens sequences (GenBank accession nos. JX083983, JX083981). B. microti sequences showed 100% identity to sequences found in I. ricinus ticks from a previous study in urban areas in Saxony, Saarland, and Bavaria (GenBank accession nos. JQ886033–JQ886058) and the human zoonotic strain Jena/Germany (GenBank accession nos. EF413181).
Sequences obtained in this study were deposited in GenBank under the following accession nos.: B. venatorum (GenBank KC007116–KC007118), B. microti (GenBank KC007119, KC007120), and B. capreoli (GenBank KC007121, KC007122).
Prevalence of Rickettsia spp. in I. ricinus
A total of 8.7% (191/2186) of questing ticks were positive for DNA of Rickettsia spp. in 2011. The following prevalences were detected in the different habitat types (Table 1): (1) Urban area, adults, 8.7% (77/889), nymphs, 5.0% (25/500), larvae, 2.2–7.5% [(2–7/93) including 2 positive pools]; (2) pasture, adults, 13.3% (30/225), nymphs, 15.7% (22/140); and (3) natural area, adults, 13.9% (11/79), nymphs, 17.5% (21/120), larvae, 2.2–10.1% [(3–14/139) including three positive pools].
There was no statistically significant difference in prevalence between months (data not shown). The prevalence in adults and nymphs was statistically significantly higher than in larvae [(p<0.01); in a positive pool of larvae, one tick was assumed as positive]. The natural and the pasture habitat showed statistically significantly higher prevalence than the urban habitat (p<0.001). Study sites M3 (urban), K (pasture), and T (natural) showed a statistically significantly higher prevalence than study site R1 and M2 (p<0.01) and exclusively M3 than B [all urban sites (p<0.05)].
gltA gene
A total of 51 Rickettsia spp. positive questing tick samples were sequenced; 47 sequences (29x urban, 9x pasture, 9x natural) had 100% identity to R. helvetica (GenBank accession nos. JQ669952, HM371185) and one sequence 100% identity (nymph, urban area, site B) to R. monacensis found in I. ricinus (GenBank accession no. JX003686). Sequencing of three specific rickettsial products failed.
ompB gene
The R. monacensis sample was further analyzed with PCR targeting the ompB gene, and sequencing revealed 100% identity to R. monacensis strain IR/Munich (GenBank accession no. EF380356). Sequences obtained in this study were deposited in GenBank under the following accession nos.: R. helvetica gltA gene (GenBank KC007123–KC007126), R. monacensis gltA gene (GenBank KC007127), and ompB gene (GenBank KC137254).
Discussion
The overall Babesia spp. prevalence in questing I. ricinus ticks did not show any obvious decrease or increase in the four city parks investigated in the 4-year survey from 2009 and 2010 (Schorn et al. 2011b) to 2011 and 2012 and ranged throughout between 0% and 2.3%. Similar rates were previously detected in Germany in the middle of an urban zone (Pichon et al. 2006), but rates were higher in renatured recreational areas in a metropolitan area in Eastern Germany (Silaghi et al. 2012). The urban sites in the present study left in a more natural condition (M3, B) offer a more suitable habitat for voles and shrews and populations of large ungulates are known there. These sites showed higher prevalence of Babesia spp. in all 4 years than the more cultivated and frequented urban sites (M2, R1) with, therefore, presumptive lower appearance of small mammals such as voles and shrews (Dickman and Doncaster 1987) and no occurrence of big mammals like roe deer (Schorn et al. 2011b).
In the present study, the highest prevalence of Babesia spp. in questing I. ricinus ticks was found in the natural area with similar occurrence of B. microti strain Jena/Germany and B. venatorum and less B. capreoli prevalence. Furthermore, 2 of 16 pools of larvae were detected positive for B. venatorum in 2012 in the natural habitat. This Babesia spp. prevalence is lower than in natural or renatured habitats in central Germany (Franke et al. 2010, Hildebrandt et al. 2010, Hildebrandt et al. 2011, Silaghi et al. 2012) or in a previously investigated forest area in Bavaria (Silaghi et al. 2012). Comparing the tick abundance of the investigated study sites and habitats (Overzier et al. 2013a, Schorn et al. 2011a, b), there was no obvious correlation between tick density and Babesia spp. prevalence. Interestingly R1, the urban study site with the highest tick density/100 m2 out of the investigated study sites from 2010–2012 (35, 97, and 51, respectively), showed the lowest or no Babesia spp. prevalence, respectively (Schorn et al. 2011a, b, Overzier et al. 2013a). The most frequently detected Babesia species in the urban area (sites M3, B) and on the pasture (K) was B. venatorum. It was only detected in those urban study sites in a more natural condition where wild ungulates exist (M3, B), whereas it was not detected in more frequented and cultivated inner city parks without large ungulate species (M2, R1) and probably less frequent appearance of small mammals as voles and shrews. This result coincides with the findings of B. venatorum (Babesia sp. EU1) during 2009 and 2010 in these four city parks (Schorn et al. 2011b). Only 1 out of 6739 I. ricinus ticks investigated in total in these four city parks over 4 years was B. microti positive, resulting in a prevalence of 0.015%. The appearance of feeding hosts for I. ricinus such as voles and shrews, which serve as natural reservoir host for B. microti (Duh et al. 2003, Karbowiak 2004, Siński et al. 2006), and roe deer, which harbors B. capreoli and B. venatorum (Babesia sp. EU1) (Duh et al. 2005, Bonnet et al. 2007a, Malandrin et al. 2010), may offer an explanation for this higher prevalence in ticks in the natural forest compared to the urban area. B. microti has been detected in Europe, for example, in the bank vole (Myodes glareolus) (Duh et al. 2003, Beck et al. 2011, Tadin et al. 2012), the common shrew (Sorex araneus) (Bown et al. 2011), and the field vole (Microtus agrestis) (Bown et al. 2011), which are suggested as reservoir hosts, but also in the yellow-necked mouse (Apodemus flavicollis) (Duh et al. 2003, Sinski et al. 2006, Beck et al. 2011, Tadin et al. 2012). In Germany, B. microti has been detected previously in a striped field mouse (Apodemus agrarius) (Silaghi et al. 2012) and with a prevalence of 1.6% in rodents of the family Arviculidae (Hartelt et al. 2008). Bank voles are most abundant in woodlands, whereas field vole and the common shrew achieve highest density in long grass and scrub (Dickman and Doncaster 1987). Both bank voles and yellow-necked mice are common in the investigated natural area (Obiegala et al., unpublished). Roe deer investigated in the natural area during 2010 and 2011 showed 89.5% positivity for Babesia spp. (B. capreoli and Babesia venatorum) (Overzier et al., 2013b).
Similar to Babesia spp., the overall prevalence of Rickettsia spp. in the urban area in 2011 was comparable to prevalence in the same study sites in 2009 (Schorn et al. 2011b). In the present study, the significantly highest prevalence for Rickettsia spp. was found in the natural area and on the pasture. The prevalence in the natural area was similar to that previously found in a geographically close forest area (site W in Silaghi et al. 2008). The lowest prevalence for Rickettsia spp. was also detected in the two more cultivated urban sites. No correlation between tick abundance and Rickettsia spp. prevalence was observed (Schorn et al. 2011a, b, Overzier et al. 2013a). The results confirm observations from another previous study where significantly less Rickettsia spp. were detected in city parks compared to forest areas (Silaghi et al. 2008). Nonetheless, even in the smallest urban study site R1, prevalence of Rickettsia spp. of up to 5.4% was detected. Other animals such as birds, which are multitudinous in study site R1 (Stadt Regensburg Umweltreferat-Gartenamt 1996), should be considered as reservoir hosts for Rickettsia spp. in urban areas. Previous studies negated (Stańczak et al. 2009) or approved (Santos-Silva et al. 2006) the reservoir potential of birds for Rickettsia spp. However, because Rickettsia spp. are not only transstadially, but also transovarially, transmitted in ticks (5 of 54 pools of questing larvae were found positive in the present study), I. ricinus should be considered as important reservoir host (Socolovschi et al. 2009).
Conclusion
For both investigated pathogens (Babesia spp. and Rickettsia spp.), the prevalence was higher in questing I. ricinus ticks collected in the natural forest area compared to the urban area. In the urban area, the prevalence was higher in more natural than more cultivated city parks. Regarding the longitudinal study, only little year-to-year variation in the occurrence of Babesia spp. and Rickettsia spp. was found. The difference in prevalence of the two pathogens most probably depends on the habitat structure and therefore the appearance or absence of diverse reservoir hosts. The potential reservoir competence of vertebrate hosts needs to be investigated in controlled settings, because the vertebrate host plays a crucial role in the life cycle of ticks and tick-transmitted pathogens and effective control measures cannot be implemented without this basic knowledge.
Footnotes
Acknowledgments
The authors address special thanks to Tim Tiedemann for assistance in laboratory work. This study was partially funded by EU grant FP7-261504 EDENext and is catalogued by the EDENext Steering Committee as EDENext083 (
Author Disclosure Statement
The authors declare that they have no conflict of interest.