
Abstract
The presence of bartonellae in invasive rats (Rattus rattus) and indigenous rodents (Arvicanthis niloticus and Cricetomys gambianus) from two districts in Uganda, Arua and Zombo, was examined by PCR detection and culture. Blood from a total of 228 R. rattus, 31 A. niloticus, and 5 C. gambianus was screened using genus-specific primers targeting the 16S–23S intergenic spacer region. Furthermore, rodent blood was plated on brain heart infusion blood agar, and isolates were verified as Bartonella species using citrate synthase gene- (gltA) specific primers. One hundred and four fleas recovered from R. rattus were also tested for the presence of Bartonella species using the same gltA primer set. An overall prevalence of 1.3% (three of 228) was obtained in R. rattus, whereas 61.3% of 31 A. niloticus and 60% of five C. gambianus were positive for the presence of Bartonella species. Genotypes related to Bartonella elizabethae, a known zoonotic pathogen, were detected in three R. rattus and one C. gambianus. Bartonella strains, similar to bacteria detected in indigenous rodents from other African countries, were isolated from the blood of A. niloticus. Bartonellae, similar to bacteria initially cultured from Ornithodorus sonrai (soft tick) from Senegal, were found in two C. gambianus. Interestingly, bartonellae detected in fleas from invasive rats were similar to bacteria identified in indigenous rodents and not their rat hosts, with an overall prevalence of 6.7%. These results suggest that if fleas are competent vectors of these bartonellae, humans residing in these two districts of Uganda are potentially at greater risk for exposure to Bartonella species from native rodents than from invasive rats. The low prevalence of bartonellae in R. rattus was quite surprising, in contrast, to the detection of these organisms in a large percentage of Rattus species from other geographical areas. A possible reason for this disparity is discussed.
Introduction
N
Bartonella species are alpha-proteobacteria related to Brucella, Agrobacterium, and Rhizobium species (Chomel et al. 2004). They are facultative intracellular organisms that invade red blood cells, often leading to long-lasting intraerythrotic bacteremia, and endothelial cells, which serve as their primary site of infection (Eicher and Dehio 2012). Clinical disease varies, ranging from mild, flu-like symptoms to more severe manifestations, such as endocarditis, myocarditis, peliosis hepatitis, and other life-threatening illnesses (Breitschwerdt et al. 2010). Furthermore, it appears that arthropod vectors may be involved in the transmission of Bartonella species to mammalian hosts, either by scarification of the skin with infected arthropod fecal material or through the bite of a blood-feeding arthropod (Billeter et al. 2008a). Fleas, lice, sandflies, and ticks have been documented experimentally to transmit a limited number of these organisms; however, Bartonella DNA has also been detected in these and other arthropods, including keds, biting flies, and mites, suggesting their potential role in the transmission of Bartonella species (Billeter et al. 2008a, Reis et al. 2011).
Rodent-borne bartonellae appear to serve as a major source of infection to humans in various locations worldwide. In Thailand, for example, B. elizabethae, B. rattimassiliensis, B. tribocorum, B. vinsonii, and B. tamiae were detected in blood clots of febrile patients and 71% of these patients reported rat exposure 2 weeks prior to illness onset (Kosoy et al. 2010). Febrile patients in Nepal were also seroreactive to B. elizabethae, B. tamiae, and B. vinsonii subsp. arupensis, suggesting contact with rodents was the source of their illness (Myint et al. 2011). In addition, rodent-associated species were suggested to be a cause of febrile illness in patients from the southwestern United States (Iralu et al. 2006). From 114 individuals, five were shown to be seroreactive against a bartonella isolated from the white-throated woodrat, Neotoma albigula, though this rodent was not necessarily the source of the infection (Iralu et al. 2006). A large percentage of intravenous drug users (IDU) from New York City, New York (46% of 205 individuals), Baltimore, MD (33% of 630 individuals), and Stockholm, Sweden (39% of 50 individuals) were observed to be seroreactive to B. elizabethae (Comer et al. 1996, Comer et al. 2001, McGill et al. 2003). Infection with rodent-borne bartonellae appears to be an issue in homeless populations, as well. Approximately 20.8% (n=48) of homeless men in Stockholm, Sweden, and 6.5% (n=200) of homeless individuals from Los Angeles, CA, harbored antibodies against B. elizabethae (Smith et al. 2002, Ehrenborg et al. 2008). The mechanism of transmission in these populations is unclear, but likely occurs through direct contact with rodents and their ectoparasites or via needle sharing, as in the case of IDUs.
The aim of the current study was to determine the prevalence of Bartonella species in invasive rats and indigenous rodent species in two districts of Uganda. It was hypothesized that individuals residing in this area may be at risk for Bartonella infection due to close contact with rodents and their fleas. Surprisingly, we found a very low prevalence in invasive rats, only three (1.3%) of 228 R. rattus harbored detectable Bartonella species, whereas ∼60% of native rodents were positive for the presence of Bartonella species either by culture and/or PCR analysis. The data presented in this study demonstrate that, if Bartonella spillover occurs, it is more likely from contact with indigenous than invasive rodents.
Materials and Methods
Rodent and flea collections
Blood and ectoparasite samples from rats were collected from control animals in a previously described study to determine the efficacy of indoor residual spraying for controlling fleas (Siphonaptera) on commensal rats in a plague-endemic region of northwest Uganda (Borchert et al. 2012). R. rattus were periodically trapped in villages residing in Arua and Zombo districts. Rodent capture/processing and rodent blood/flea collections were described previously (Borchert et al. 2012). For this study, rodents trapped in nontreated sites (Arua villages of Tabanzu and Kaza; Zombo villages of Gbalia, Jupakonja, and Oyaragada) and their fleas were examined (Table 1). A. niloticus and C. gambianus were trapped in Arua Town in the Arua district (Table 1); however, fleas from peridomestic rodents were not collected.
Blood (whole blood or blood clot) was collected from commensal, Rattus rattus, and peridomestic, Arvicanthis niloticus and Cricetomys gambianus, rodents from two districts in Uganda, Arua and Zombo. Additionally, fleas were recovered from R. rattus. A total of 264 rodents and 104 fleas were examined for the presence of Bartonella species.
Whole blood was obtained from rodents either in EDTA tubes or in 2.0-mL microcentrifuge tubes. Fleas were removed from hosts and placed in microcentrifuge tubes containing 70% ethanol. Rodents and fleas were subsequently identified to species using taxonomic keys (Hopkins 1947, Haselbarth 1966, Smit 1973, Delaney 1975). All samples were shipped on dry ice to the Bartonella Laboratory, Centers for Disease Control and Prevention, Fort Collins, Colorado, for analysis.
Sample processing and screening
DNA was extracted from 100 μL of whole blood or blood clot following the Qiagen QIAamp tissue kit protocol (Qiagen, Valencia, CA) per the manufacturer's instructions. Individual fleas were triturated using a sterile needle, and DNA was extracted using the same tissue protocol. Negative extraction controls, specifically reagent solutions only, were included with each set of extractions. A total of 228 R. rattus, 31 A. niloticus, five C. gambianus, and 104 fleas (21 X. cheopis, 70 X. brasiliensis, 12 D. lypusus, and one A. torvus) were examined.
For culture, 100 μL of whole blood or blood clot was mixed with brain heart infusion (BHI) broth (CDC, Atlanta, GA) containing 10% fungizone at a dilution of 1:2 for whole blood or 1:4 for blood clot. One hundred microliters of the whole blood suspension or 200 μL of the blood clot suspension was aliquoted onto a BHI agar plate containing 10% rabbit blood (CDC). Plates were placed at 35°C, 5% CO2 and examined weekly for up to 4 weeks. Any Bartonella-like colonies were subsequently subpassaged onto fresh agar plates, and DNA was extracted from colonies using the same protocol described above.
DNA extracted from blood was screened by conventional PCR using primers targeting the Bartonella 16S–23S intergenic spacer region (ITS), as described previously (Billeter et al. 2008b). Bacterial cultures were confirmed as Bartonella species, and fleas were examined for the presence of bartonellae DNA using citrate synthase-specific primers (gltA) and the same protocol described by Billeter et al. (2011). Amplicons were purified using the QIAquick PCR purification kit (Qiagen) and sequenced using an Applied Biosystems Model 3130 genetic analyzer (Applied Biosystems, Foster City, CA). DNA sequences were analyzed using the Lasergene version 8 sequence analysis software (DNASTAR, Madison, WI). For gltA sequences, sequences were shortened to ∼379 bp, if possible, to allow for further phylogenetic analysis (La Scola et al. 2003). gltA sequences obtained in this study were considered similar to validated Bartonella species if similarity over the 379-bp fragment was ≥96% (La Scola et al. 2003). The Clustal W program in MegAlign (Lasergene) was used to compare sequences obtained from this study to Bartonella sequences in GenBank.
Nucleotide sequence accession numbers
Sequences representative of the gltA genotypes obtained from this study were deposited in GenBank under accession numbers JX428744–JX428750 (bartonellae cultured from rodent blood) and JX428759–JX428765 (genotypes detected in fleas).
Results
Bacteria detected in the blood
Of the 264 rodents examined, two R. rattus (one rat from Tabanzu and another from Jupakonja villages), 11 A. niloticus, and three C. gambianus were PCR positive when blood was screened directly using ITS primers. The bacteria detected in both R. rattus (GenBank accession nos. JX428751 and JX428752) was 96.0% similar to Bartonella sp. RN28BJ (GenBank accession no. EF213776), related to B. elizabethae, found in R. norvegicus from Beijing, China. The ITS sequences detected in peridomestic rodents was 89–95% similar to sequences available in GenBank, but over a limited query coverage (32–82%). Therefore, genetic relatedness could not be determined using these sequences. ITS sequences obtained from indigenous rodents, containing six distinct genotypes, have been deposited in GenBank under accession numbers JX428753–JX428758.
Bacteria isolated from blood
Two R. rattus (the one from Tabanzu village and an additional from Jupakonja) harbored viable organisms (Table 2 and Fig. 1). A single genotype was detected in the rat from Tabanzu (genotype 1, GenBank accession no. JX428744), and this genotype was 97.6%, 97.1%, and 96.2% similar to Bartonella sp. PD5700t found in Praomys delectorum from Tanzania (GenBank accession no., FJ851115), Bartonella sp. RRB046N identified in R. rattus from Nepal (GenBank accession no. GU143527), and B. elizabethae (GenBank accession no. U28072), respectively. A single genotype was detected in the Jupakonja R. rattus isolates (genotype 2, GenBank accession no. JX428745) and was 99.5%, 98.9%, and 96.4% identical to Bartonella sp. Pd5700t, RRB046N, and B. elizabethae, respectively.
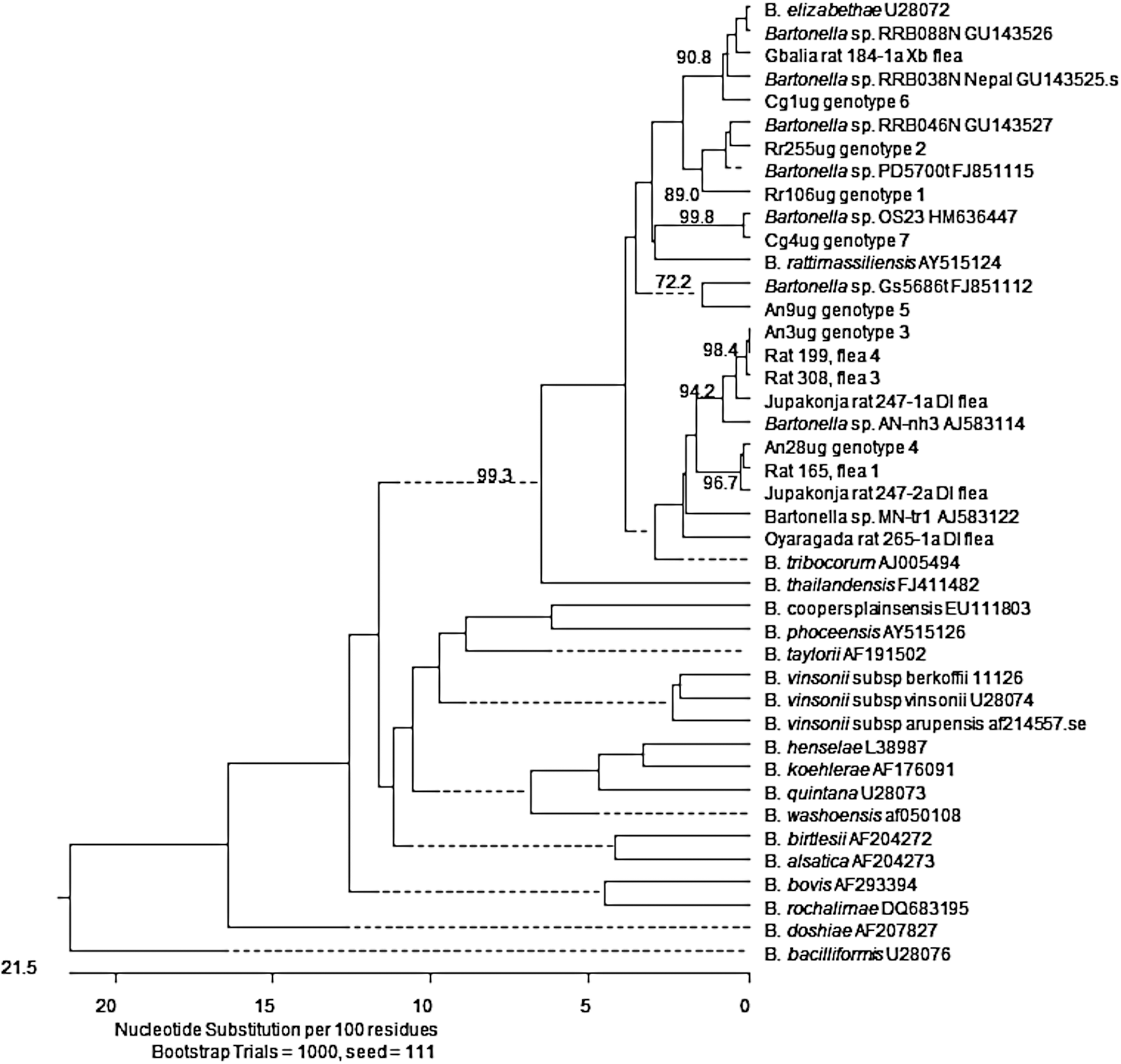
Phylogenetic classification of Bartonella genotypes detected in Rattus rattus and their fleas, Arvicanthis niloticus, and Cricetomys gambianus trapped in two districts of Uganda based upon a partial citrate synthase gene gltA. The phylogram was constructed by the neighbor-joining method with the Kimura-2 parameter model. Only bootstrap values above 70% obtained from 1000 replicates are given. gltA sequences from this study are represented by GenBank accession numbers JX428744–JX428750 from rodents and JX428759–JX428765 from R. rattus fleas.
Rattus rattus, Arvicanthis niloticus, and Cricetomys gambianus, were screened for the presence of Bartonella species using PCR and culture methods. Blood was plated onto agar plates and isolates were identified using Bartonella citrate synthase gene, gltA, primers. Isolates from 10 of 17 A. niloticus were characterized. Isolates obtained in this study were considered similar to validated Bartonella species if similarity over the 379 bp gltA fragment was ≥96% (La Scola et al. 2003).
A. niloticus #28 was infected with two Bartonella species: genotypes 3 and 4. All remaining rodents were infected with a single bartonella.
From indigenous rodents, Bartonella species were isolated from 17 A. niloticus (eight rodents were bacteremic but PCR negative; two were PCR positive but not bacteremic, with a total of 19 PCR and/or culture positives) and three C. gambianus. Sequence confirmation was obtained for 10 of the 17 A. niloticus samples. In most cases, a single genotype was isolated from these rodents, excluding A. niloticus no. 28, which was infected with two bartonellae genotypes (Table 2). Eight of the 10 A. niloticus (100% similarity among isolates, genotype 3, GenBank accession no. JX428746) harbored a bacterium related to Bartonella sp. AN-nh3 (GenBank accession no. AJ583114), originally isolated from Micaelamys namaquensis in South Africa, with 98.0% similarity (Fig. 1). In addition, A. niloticus no. 17 and no. 28 (genotype 4, GenBank accession no. JX428747) also were infected with a genotype related to Bartonella sp. MN-tr1 (GenBank accession no. AJ583122), found in the blood of Mastomys natalensis also from South Africa, with 96.9% sequence similarity. In a single A. niloticus (genotype 5, GenBank accession no. JX428748), a genotype 97.3% similar related to an uncultured Bartonella sp. detected in a Grammoys sp. from Tanzania (GenBank accession no. FJ851112), was isolated. A genotype, related to B. elizabethae with 98.5% similarity, was isolated from a C. gambianus (genotype 6, GenBank accession no. JX428749), while the remaining two C. gambianus (genotype 7, GenBank accession no. JX428750) were infected with a bartonella initially isolated from a soft tick, Ornithodorus sonrai, from Senegal (GenBank accession no. HM636447) with 99.7% sequence similarity.
Bacteria detected in fleas from invasive rats
Of 104 fleas examined, 6.7% (seven total) harbored detectable Bartonella DNA: 6 D. lypusus and 1 X. brasiliensis were PCR positive (Table 3). Two D. lypusus (R. rattus 199, flea 4 and R. rattus 308, flea 3), recovered from a rat in the villages of Kaza and Tabanzu, harbored bacteria that were 100% identical to the Bartonella sp. genotype 3 and 98.0% similar to Bartonella sp. AN-nh3 (Fig. 1). An Oyaragada village D. lypusus (R. rattus 265, flea 1) was also infected with a bartonella (GenBank accession no. JX428764) that was 96.2% similar to Bartonella sp. AN-nh3. A D. lypusus (R. rattus 247, flea 1), recovered from the Jupakonja village, harbored a sequence that was 99.1% identical to Bartonella sp. AN-nh3 (GenBank accession no. JX428761). Additionally, a second D. lypusus flea (R. rattus 247, flea 2), from the same Jupakonja rat, was infected with a Bartonella sp. (GenBank accession no. JX428762) that was 99.7% similar Bartonella sp. genotype 4 and 96.5% similar to Bartonella sp. MN-tr1. A Gbalia village D. lypusus flea (R. rattus 165, flea 1) also harbored a bartonella that was 100% identical to Bartonella sp. genotype 4 and 97.0% similar to Bartonella sp. MN-tr1. B. elizabethae DNA was detected in the single X. brasiliensis–positive flea (R. rattus 184, flea 1; GenBank accession no. JX428760 with 99.3% sequence similarity) recovered from a R. rattus from the village of Gbalia.
A total of 104 fleas (21 Xenopsylla cheopis, 70 X. brasiliensis, 12 Dinopsyllus lypusus, and 1 Afristivalius torvus), collected from Rattus rattus, were screened using Bartonella citrate synthase gene, gltA, primers; seven fleas harbored detectable Bartonella species. Flea sequences obtained in this study were considered similar to validated Bartonella species if similarity over the 317-bp gltA fragment was ≥96% (La Scola et al. 2003).
Discussion
Surprisingly, a very low Bartonella prevalence was found in R. rattus populations from the two districts, Arua and Zombo, in Uganda; only three of 228 rats examined were positive. This contrasts with numerous studies demonstrating a high prevalence of bartonellae infection among Rattus rats in Bangladesh, China, Israel, Nepal, Thailand, and the United States (Ellis et al. 1999, Bai et al. 2007, Inoue et al. 2008, Bai et al. 2009, Harrus et al. 2009, Saisongkorh et al. 2009, Gundi et al. 2010). We speculate that the relatively low prevalence of infection may be due to the fact that Rattus rats were a historically “recent” introduction into this area of the West Nile region. Only a single specimen was reported despite intensive investigations in 1937–1938 (Amatre et al. 2009) with rat numbers increasing in the 1940s–1950s (Hopkins 1949). Movement of R. rattus throughout the world is generally associated with the dispersal of humans and their cargo via ships and other forms of transportation (Aplin et al. 2011). As this region was maintained as a “closed area” (i.e., outsiders were excluded and movement of crops limited) prior to 1914 when the region became a British protectorate (Borchert et al. 2007), this likely hampered the relocation of R. rattus and thereby limited the introduction of Bartonella-infected rats. Further investigations screening Rattus species from different sites throughout the West Nile region are warranted to determine if similarly low bartonellae prevalence is present in those rodent populations.
Unlike invasive rats, a large percentage of A. niloticus (61.3%) and C. gambianus (60%) were infected with Bartonella species. The majority of A. niloticus harbored bartonellae similar to those previously identified in Micaelamys namaquensis, the Namaqua Rock Rat, from the Free State province in South Africa (Pretorius et al. 2004). Isolates from two A. niloticus were similar to a species found in M. natalensis from the same South African study (Pretorius et al. 2004), and a single A. niloticus was infected with a genotype related to a Bartonella detected in the spleen of a Grammoys sp. from Tanzania (Gundi et al. 2012). Furthermore, two C. gambianus were infected with bacteria similar to a bartonella isolated from the soft tick O. sonrai from Senegal (Mediannikov et al., unpublished; GenBank no. HM636447). The results from this study demonstrated a lack of host specificity because similar Bartonella genotypes were isolated from a diverse host range and broad geographical area. This would suggest that transmission of these organisms may occur by shared or common ectoparasites, such as one or more species of flea or tick, found distributed throughout the African continent. Whether these Bartonella species can infect and cause clinical disease in humans and other animals remains to be determined, as well.
B. elizabethae DNA was detected and/or isolated in the blood of a C. gambianus and three R. rattus in this study. As described previously, B. elizabethae is typically associated with commensal rats (Rattus spp.) and is a suggested cause of infective endocarditis in humans and domestic dogs (Daly et al. 1993, Mexas et al. 2002) and is associated with febrile illness in Thai patients (Kosoy et al. 2010). The zoonotic potential of this organism raises concerns because individuals residing in these areas are frequently exposed to rodents and their ectoparasites, which greatly increases the opportunity for disease transmission. Although the infection prevalence for persons residing in Uganda is unknown, a study performed by screening patients from the Democratic Republic of Congo found that 4.5% of 155 individuals were Bartonella seroreactive (Laudisoit et al. 2011). None of these patients harbored antibodies to rodent-borne bartonellae, such as B. elizabethae, B. vinsonii subsp. vinsonii, or B. vinsonii subsp. arupensis, but a similar seroprevalence rate may not be found in this region of Uganda. Interestingly, the Gambian pouched rat was found infected with B. elizabethae; this fact is important from public health perspective because rats of this species commonly are found in fields and gardens near villages and are commonly hunted and prized by local cultures in the West Nile region. As with other blood-borne pathogens, the public must be made aware that Bartonella may be transmitted through contact with contaminated meat or blood products, as was the case for an elderly patient who became infected with B. alsatica while butchering rabbits (Angelakis et al. 2008).
Fleas recovered from invasive rats harbored Bartonella DNA similar or identical to species identified in native rodent species and not their natural hosts. Earlier surveillance in the two Ugandan districts found that D. lypusus were typically associated with A. niloticus or M. natalensis, X. brasiliensis was commonly found on A. niloticus or R. rattus, and X. cheopis were encountered most frequently on M. natalensis or R. rattus (Amatre et al. 2009). Our results would suggest that flea sharing among these rodents is quite common and would therefore explain the detection of bartonellae related to M. namaquensis and M. natalensis genotypes in R. rattus–associated fleas. It must be stipulated, however, that experimental transmission studies to determine arthropod vector competency for most Bartonella species are lacking.
Despite the low Bartonella prevalence in invasive rats, our data confirm that infection rates are quite high in certain indigenous rodent populations. Furthermore, flea sharing does occur between invasive rats and native rodents in Uganda. Further work is required to identify whether Bartonella species do infect people residing in Uganda, what role these bacteria may have in clinical disease, and whether rodent fleas are competent vectors of Bartonella species.
Footnotes
Author Disclosure Statement
No competing financial interests exist for any author.