
Abstract
Tick-borne diseases pose an increasingly important public health problem in Europe. Rodents are the reservoir host for many tick-transmitted pathogens, including Anaplasma phagocytophilum and Babesia microti, which can cause human granulocytic anaplasmosis and babesiosis, respectively. To estimate the presence of these pathogens in rodents in Finland, we examined blood samples from 151 bank voles (Myodes glareolus) and demonstrate, for the first time, that A. phagocytophilum and B. microti commonly infect bank voles (in 22% and 40% of animals, respectively) in Finland. Sequence analysis of a fragment of 18S rRNA showed that the B. microti strain isolated was identical to the Munich strain, which is considered to be nonzoonotic. The A. phagocytophilum strain (based on a fragment of the msp4 gene) was identical to one found earlier in rodents in the United Kingdom that is transmitted by the tick Ixodes trianguliceps, all the life stages of which feed on small mammals. The infection probability of B. microti in the bank voles was the greater the older the individual was, and males were more often infected than females. A. phagocytophilum infection probability first increased and then decreased with the age of individual without any difference between sexes. While these pathogens presumably pose a limited zoonotic risk to humans in Finland, they might have important interactions with other rodent pathogens and therefore affect infection dynamics of, for example, zoonotic pathogens.
Introduction
T
Babesia microti is the main causative agent for human babesiosis in the United States (Gray et al. 2010). Although there is only one confirmed human infection by B. microti in Europe (Hildebrandt et al. 2007), because most cases are caused by B. divergens (Gray et al. 2010), serological studies indicate that human B. microti infections are more common than previously recognized (Hunfeld et al. 2002). B. microti is widely distributed across the Northern Hemisphere (Goethert and Telford 2003), and at least two B. microti strains (Munich and Jena), which may differ in terms of their association with human disease, are found in Europe (Pieniazek et al. 2006, Siński et al. 2006, Bown et al. 2008, Beck et al. 2011, Katargina et al. 2011).
In Europe, some 70 confirmed HGA cases have been reported to date (Dumler et al. 2005, Edouard et al. 2012). However, seroepidemiological studies have revealed high (up to 21%) A. phagocytophilum antibody prevalences in some areas in Europe (Dumler et al. 2005), suggesting that asymptomatic human infections might be common (Doudier et al. 2010). A. phagocytophilum also has indirect implications for humans because it causes, for example, a tick-borne fever in livestock and febrile fevers in companion animals (cats and dogs) (Rar and Golovljova 2011). Many wildlife mammalian species, such as rodents, shrews, and cervids, are naturally infected with A. phagocytophilum (Rar and Golovljova 2011). Different genetic variants of A. phagocytophilum circulate in different tick and mammalian species (Bown et al. 2008), and these strains may differ in their pathogenicity to humans (Doudier et al. 2010). The distribution of A. phagocytophilum is worldwide (Rar and Golovljova 2011).
Wild rodents and shrews are important reservoir hosts for many tick-borne pathogens, and therefore they have a key role in the natural circulation of tick-borne infections, including A. phagocytophilum and B. microti (Goethert and Telford 2003, Bown et al. 2008, Bown et al. 2011). In the rodent reservoirs, A. phagocytophilum infection is transient, lasting for 1–2 months, whereas B. microti infection is chronic (Bown et al. 2003, Bown et al. 2008). The transmission of both A. phagocytophilum and B. microti is transstadial, i.e., an uninfected tick (larvae or nymph) becomes infected while feeding on an infectious reservoir host that itself had become infected while feeding an infected tick (nymph or adult) (Gray et al. 2002, Woldehivet 2006, Hunfeld et al. 2008). The primary vectors of A. phagocytophilum and B. microti are ticks from the genus Ixodes (Gray et al. 2002, Woldehivet 2006, Hunfeld et al. 2008).
Here we quantify the presence and prevalence of A. phagocytophilum and B. microti in Finland in the bank vole (Myodes glareolus) and identify the pathogen strains that are circulating in the host population. We find that A. phagocytophilum and B. microti infection prevalence in bank voles is high (22% and 40%, respectively), and we provide the first evidence that these pathogens are circulating in Finnish rodents.
Materials and Methods
Rodent trapping was carried out in July, 2008, as a part of an ongoing long-term study on bank vole (M. glareolus) populations in the Konnevesi area in central Finland (62°83’79” N, 26°82’09” E) (for details of the study sites and trapping procedures, see Kallio et al. 2009). The bank vole dominates the small rodent community in the study sites (our unpublished data). All captured bank voles (63 females and 88 males) were taken to the laboratory where their body mass, head width, and sex were recorded and a blood sample was taken from the retro-orbital sinus with capillary tubes (Haematocrit Capillaries, Hirschmann Laborgeräte, Germany). The presence of ticks was not examined. The samples were placed on dry ice immediately after collecting and stored at −80°C until use.
Presence or absence of pathogens was assessed using PCR-based assays. Briefly, total DNA was extracted from bank vole blood using alkaline digestion (for details, see Bown et al. 2003). Negative controls (one per every four samples) were included in the DNA extraction and in the PCR assays, and none of the negative controls were found positive in the PCR assays. Detection of A. phagocytophilum used a nested PCR-assay that targets the 16S rRNA gene (primers in Massung et al. 1998; PCR conditions are described by Bown et al. 2006). The positive control was a DNA extract derived from a strain of A. phagocytophilum isolated from a sheep (Ovis aries) in the United Kingdom. To quantify the strain variation, six randomly chosen positive samples were screened using a separate nested PCR that targets a fragment of the msp4 gene, with thermal cycling conditions and primers described by Bown et al. (2007); amplicons were sequenced. B. microti was detected using an Apicomplexa-specific nested PCR that targets a fragment of the 18S rRNA gene (Simpson et al. 2005). The positive control used was B. microti Munich strain. Because this method is not B. microti specific, all putative positive samples were sequenced to separate B. microti from Hepatozoon sp. infections. Sequence identify was determined by BLAST search (Altschul et al. 1990) against the National Center for Biotechnology Information (NCBI) Nucleotide database. Only samples that were confirmed to be infected by B. microti on the basis of the sequence data were considered as positive results.
We analyzed separately the probability of being infected (binary response variable) with A. phagocytophilum and B. microti using a generalized linear mixed model (GLMM) approach with a logit link function and binomial distribution. The explanatory variables assessed in the full model were gender, head width, and head width squared and whether the individual was co-infected with the other parasite. Body size is often used as a proxy for age in wild rodents. In central Finland, for example, bank vole head width is approximately on average 8.1–8.4 mm at birth, 11.6–12.0 mm at weaning (∼3 weeks age), 12.7 mm and 13.4 mm at the age of ∼2 and ∼5 months, respectively, and 13.8–14.0 at the age of 10 months (Koskela et al. 1998, Oksanen et al. 2001, Kallio et al. 2006, our unpublished data). Models were fitted using the Laplace approximation method (lmer function in lme4 package in R software, available under GNU license at
Results
Out of the 151 bank voles captured in July, 2008, 33 individuals (21.9%; 95% confidence interval [CI] 15.3–28.5) were infected with A. phagocytophilum, 60 individuals (39.7%; 95% CI 31.9–47.5) were infected with B. microti, and 11 individuals (7.3%; 95% CI 3.1–11.4) were infected with both pathogens. The probability of a bank vole being infected with A. phagocytophilum peaked at intermediate ages (i.e., head width of ∼12.5–13.5 mm, which corresponds approximately the age of 2–5 months; see Materials and Methods) (Fig. 1, Table 1). Neither host sex nor B. microti infection status had a significant effect on A. phagocytophilum infection probability. B. microti infection probability increased with age (increasing head width) in both sexes with males being more likely to be infected than females (Fig. 2, Table 2). A. phagocytophilum infection did not influence the probability of being infected with B. microti.
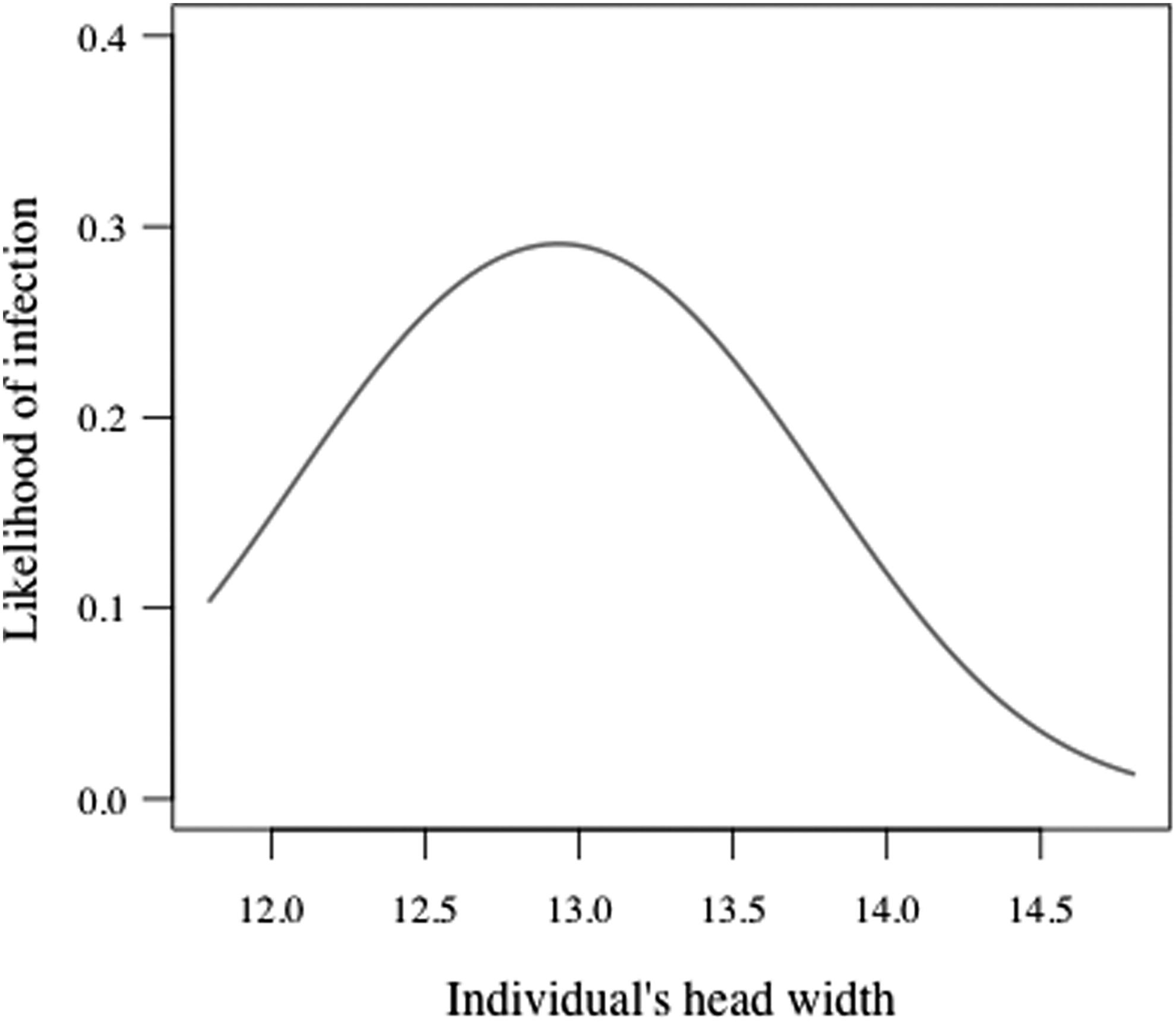
Predicted probability of being infected with A. phagocytophilum in relation to individual's head width (mm).
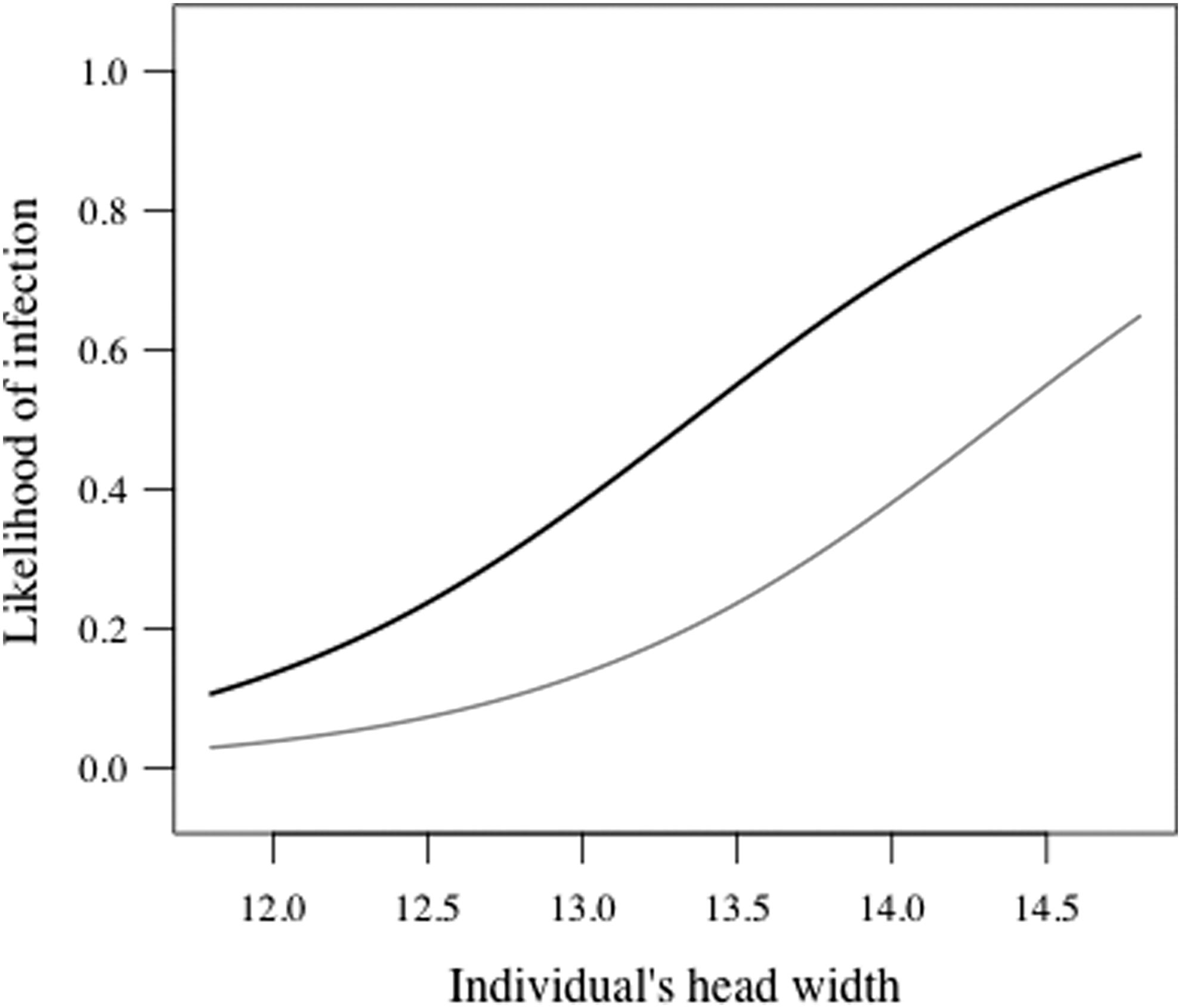
Predicted probability of being infected with B. microti in relation to individual's head width (mm) and sex. (Black line, top) male; (grey line, bottom) female.
σ2 is the variance attributable to random effect.
sd, standard deviation of σ2; SE, standard error.
σ2, the variance attributable to random effect.
sd, standard deviation of σ2; SE, standard error.
Analysis of the msp4 fragment showed the strain of A. phagocytophilum to be identical to that previously reported in field voles in the United Kingdom (GenBank accession no. FJ469653) (Bown et al. 2007). All 60 sequences of 18S rRNA fragment from putative positive B. microti–infected animals were identical to the Munich strain of B. microti (GenBank accession no. AB071177).
Discussion
In this study we present the first evidence that B. microti and A. phagocytophilum pathogens are present in wild rodents in Finland. A. phagocytophilum is known to cause tick-borne fever in cattle in Finland (Tuomi 1966), and, recently, a domestic cat was diagnosed with A. phagocytophilum infection (Heikkilä et al. 2010). B. microti, in turn, has been reported in a single Ixodes persulcatus tick in western Finland (Alekseev et al. 2007). However, this is the first study examining the presence of A. phagocytophilum and B. microti in wildlife reservoir host species.
The B. microti Munich strain found in this study has been suggested not to be pathogenic to humans (Gray 2006, Siński et al. 2006), although it may be transmitted by I. ricinus ticks (Welc-Falęciak et al. 2012), which is the most important tick species in transmitting tick-borne pathogens to humans in Europe (Jaenson et al. 2012a). In the United Kingdom, however, B. microti Munich is transmitted by the nidiculous tick Ixodes trianguliceps, all life stages of which feed on rodents (Randolph 1995, Bown et al. 2008). Similarly, the A. phagocytophilum strain found here is identical to the one found in the field vole (Microtus agrestis) in the United Kingdom, which was apparently restricted to an enzootic small mammal–I. trianguliceps cycle (Bown et al. 2006, Bown et al. 2008, Bown et al. 2009). Therefore, it is not likely that the B. microti or A. phagocytophilum strains found here pose any direct risk to humans in Finland. However, this study was carried out in only one location, and only bank voles were examined. Because different strains of A. phagocytophilum may exploit different tick and mammal species supporting separate enzootic cycles (Bown et al. 2008), it is possible that other genetic variants are circulating in Finland. Moreover, B. microti strain Jena, which is considered to be zoonotic, has been found in neighboring Estonia (Katargina et al. 2011), which may suggest that this strain could be also found in Finland. Hence, further work has to widen the scope of sampling and, especially, focus on questing Ixodes ticks, which pose a risk to humans, to determine whether zoonotic A. phagocytophilum or B. microti strains exist in Finland.
Even if the A. phagocytophilum and B. microti found in bank voles do not pose a direct risk to humans in Finland, these pathogens may interact with other pathogens, within their rodent hosts, because these pathogens have significant interactions with other infections in field voles in the United Kingdom (Telfer et al. 2010). For instance, A. phagocytophilum infection increases cowpox virus and decreases Bartonella sp. and B. microti infection probabilities, whereas B. microti decreases the risk of Bartonella sp. infection while increasing the risk of A. phagocytophilum infection. These strong individual level effects are likely to be translated to infection dynamics at the population level (Telfer et al. 2010), and for zoonotic pathogens may be further reflected in human epidemics.
We do not know yet which tick species is responsible for the circulation of these pathogens in our study area, where both I. ricinus and I. trianguliceps are found (our unpublished data). In the United Kingdom, the same strains of A. phagocytophilum and B. microti are transmitted primarily by I. trianguliceps (Bown et al. 2008). The distribution of I. trianguliceps covers most of Finland (Ulmanen 1972), thus the pathogens transmitted by I. trianguliceps might also be found across Finland. Moreover, I. trianguliceps infests many small mammal species, such as bank voles (Myodes glareolus), field voles (Microtus agrestis), root voles (Microtus oeconomus), common shrews (Sorex araneus), and pygmy shrews (S. minutus) (Ulmanen 1972, Nilsson 1974, Randolph 1975, Bown et al. 2003, Bown et al. 2011), many of which are competent reservoir hosts species for a variety of tick-borne pathogens, including A. phagocytophilum and B. microti (Randolph 1995, Liz et al. 2000, Bown et al. 2003, Bown et al. 2006). Therefore, it is likely that A. phagocytophilum and B. microti are widely distributed in small mammal species in Finland.
The infection prevalences reported here are in line with earlier studies on A. phagocytophilum and B. microti in rodents from elsewhere in Europe (e.g., Liz 2002, Duh et al. 2003, Siński et al. 2006, Bown et al. 2008, Bown et al. 2011, Beck et al. 2011, Rar et al. 2011). The peak in A. phagocytophilum infection likelihood at intermediate individual age (i.e., head width) is likely to reflect the transient nature of this infection (Bown et al. 2003), whereas the constantly increasing likelihood of B. microti infection with age reflects a chronic infection (Bown et al. 2008). Although both pathogens are likely to be transmitted by the same vector species, only B. microti showed some difference in infection probability between sexes (more common among males).
Conclusion
To summarize, A. phagocytophilum and B. microti are prevalent in bank voles in central Finland. Because they are transmitted by I. trianguliceps ticks (Bown et al. 2008), whose distribution range covers most of Finland (Ulmanen 1972), we predict that these pathogens might be widespread in Finland. Although it is unlikely that B. microti or A. phagocytophilum strains found here cause any direct risk to humans in Finland, these pathogens may interact with other pathogens (Telfer et al. 2010) and therefore affect infection dynamics of, for example, zoonotic pathogens. Therefore, further studies are needed to examine the role of these B. microti and A. phagocytophilum strains in the pathogen community of Finnish rodents and to examine the presence of other, potentially zoonotic strains in Finland.
Footnotes
Acknowledgments
This research adhered to the Association for the Study of Animal Behaviour/Animal Behaviour Society Guidelines for the Use of Animals in Research, the legal requirements in Finland, and all institutional guidelines. We wish to thank H. Helle for help with fieldwork and sampling. This study was financially supported by Academy of Finland (grant nos. 250524 to E.R.K.; 218107 and 257340 to E.K.; 132190 to T.M.). We are also grateful for support provided by the Konnevesi Research Station.
Author Disclosure Statement
No competing financial interests exist.