
Abstract
Tick-borne encephalitis (TBE) is a human disease caused by a flavivirus that is spread by ticks (Ixodes ricinus). In 2011 and 2012, the highest TBE incidence ever was recorded in Sweden. It has been proposed that warmer spring temperatures result in higher survival of ticks and thus high incidence of TBE. Here, analyses were done of time series of TBE for 1976–2011 in relation to the North Atlantic Oscillation (NAO), mean summer temperatures, and yearly number of harvested European hare (Lepus europeaus), roe deer (Capreolus capreolus), and red fox (Vulpes vulpes) in the County of Stockholm, the area with most TBE cases in recent years in Sweden. The results show that the winter NAO index or winter temperature has no significant effect on the variation in wildlife numbers harvested or TBE cases over time. Mean summer temperature above 12°C had a slight effect, but a multivariate model revealed that only the numbers of European hare and red fox remained in the model and explained 64.4% of the variation in TBE cases. Ticks do not seem to be as sensitive to climate variations as anticipated, even though that summer temperature has increased by 2°C during the time period studied here. Instead, TBE cases seem to be more dependent on host population dynamics than on climate factors.
Introduction
T

Recent distribution of human tick-borne encephalitis (TBE) cases in Sweden. (From
This expansion and spatial increase is puzzling because a vaccination program for humans has been in place in Sweden against the virus for several years. Lindgren and Gustafsson (2001) suggested that the increase in the disease incidence of TBE in humans was related to events of mild winters. On the other hand, Randolph (2004, 2010) and Sumilo et al. (2007) found no climate effect on TBE in a country-based analysis in Europe. The reason for the increase in TBE in Sweden remains unknown, mainly because our limited knowledge on the role of wildlife and environmental changes and how they contribute to increased risk for infections in humans (Ostfeld and Keesing 2000, Schmidt and Ostfeld 2001, Keesing et al. 2006). Climate variations could be assumed to be decisive for the TBE dynamics because many biological, behavioral, and environmental factors for ticks are weather dependent (Epstein 2002).
Recent studies show that there is a relationship between large-scale climatic systems such as the North Atlantic Oscilliation (NAO) and El Nino Southern Oscillation (ENSO) with life history traits and population dynamics of animals and some animal-borne diseases (Post and Stenseth 1999, Murua et al. 2003). NAO determines weather patterns over Europe during winter and reflects the changes in air pressure over the North Atlantic (Hurrell 2012). The NAO is positively related to temperature, and a high NAO index corresponds to mild winters over Scandinavia, whereas a low index corresponds to cold winters. The NAO shows variability over time and could serve as a tool for analyzing the sensitivity of various living organisms to climate change (Post and Forchhammer 2002). The effect by climate is both indirect through a tropic cascade via increased primary productivity and direct through higher survival rates of animals, vectors, and disease agents (Catchpole et al. 2000, Coulson et al. 2001).
Even though the NAO climate index has increased and global temperature is increasing, few studies have demonstrated direct climate effects on disease systems and host dynamics (Lindsay and Birley 1996, Schmaljohn and Hjelle 1997, Engelthaler et al. 1999, Palo et al. 2005, Palo 2009). Greater survival rates and hence generally higher population densities of host species are expected to increase the transmission of virus, but the situation is more complex than anticipated. A recent study put forward that several mammal species and some bird species are competent hosts for ticks and the flavivirus causing TBE (Jaenson et al. 2012). According to this study, roe deer (Capreolus capreolus) are regarded as the main host for ticks, but hares (Leporidae) and small rodents (Microtinae) may be equal or more important (Tälleklint and Jaenson 1993, Laurenson et al. 2003).
Furthermore, effects by predation on the host population or regulation of the predator may have a cascading effect on the disease agent by its effect on host populations (Willmers et al. 2006). In spite of these observations, we know very little about the dynamic effects caused by multihost interactions and synergies between factors affecting transmission risk. It has been shown that disease transmission is a population-driven process with feedbacks between hosts, disease agents, and environmental factors (Anderson and May 1978, Anderson and May 1985, Altizer et al. 2006). In view of our limited knowledge about the ecological factors involved in disease transmission, it is important to develop models based on collection of new relevant data and to generate new testable hypotheses about ecological factors operating on zoonotic disease systems. Here time series of mammalian host species populations, TBE, NAO, and temperature data are combined to reveal causal relationships and to identify the role of climate variation and other driving factors for TBE transmission in Sweden.
Methods
The time series were selected for the length of the series and mammalian species known to be associated with ticks as host animals (Laurenson et al. 2003). People move through vegetation in their sport and outdoor activities and thereby encounter ticks as they move. Human TBE cases are here assumed to reflect the infected tick population. Encounters of infected ticks by humans are regarded as a random process that is in direct proportion to the infected tick population. Likewise, harvest of mammals in hunting bags is a process related directly to the population size of the hunted species. Variations in hunting efficiency provide a good estimate of the actual population density (Cattadori et al. 1999, Haydon et al. 2002). Accordingly, TBE in humans would also reflect infected ticks in relation to host abundance and an indirect estimate of transmission efficiency Ro.
Time series covering hunting statistics for roe deer, European hare, and red fox for 1976–2011 and for the County of Stockholm were provided from the hunting statistics held by the Swedish Association for Hunting and Wildlife Management. Monthly temperature data was provided from the Stockholm observatory corrected for urban influence. Temperature means for the months November–March and months April–October were used as well as the NAO index for the same periods. The former period is here denoted winter season and the latter summer season for both temperature series and the NAO index time series. NAO data were downloaded from Hurrell (2012). Data series of TBE cases covering 1976–2012 were obtained from the Swedish Center for Disease Prevention (SMI). To conform to parametric statistics, data were log transformed if necessary and statistical analyses were done in XLSTAT version 2013.1.01.
Results
Climate and temperature time series
The winter NAO climate index showed a positive relationship with winter season temperatures at Stockholm observatory (y=0.4584x+0.6418; R 2=0.4755, p<0.001), but no such relationship was found between the winter season NAO and mean temperatures for the summer season. There was no significant relationship between the winter NAO index and summer NAO index, which implies independence in seasonal climate; the variation in summer NAO was lower than for winter NAO. The mean summer temperature measured at Stockholm observatory has increased from 1976 to present with about 2°C (y=0.058x+10.53; r 2=0.54, p<0.001).
TBE and climate factors
Human TBE cases show an exponential increase over time, with an all-time high in 2011 and 2012 with 423 and 320 cases, respectively. No significant difference in TBE cases was found between the high winter NAO and low winter NAO index (t-test, p=0.892, degrees of freedom [df ]=32). There is a relationship between number of TBE cases and summer temperature, as shown in Figure 2, and temperature explains 19.4% of the variation in TBE cases as a single explanatory variable. The increase in TBE cases from 1976 is particularly evident over a break point for the increase at temperatures with a mean exceeding 12°C (t-test, p<0.0001, df=34). Thus, summer temperature could be one important factor facilitating increase in TBE cases in Sweden.
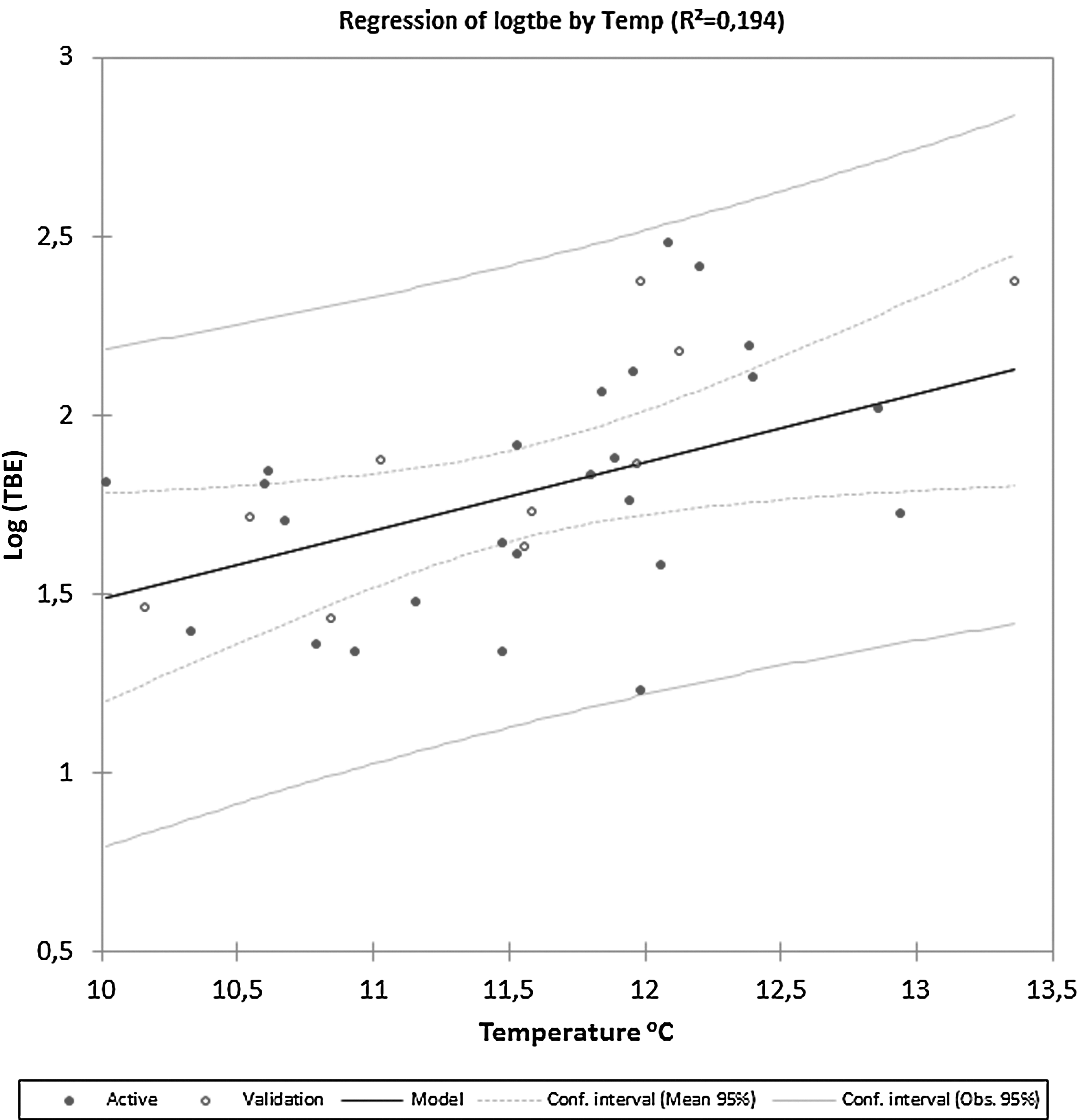
Relationship between mean summer temperatures (April–October) and total cases of tick-borne encephalitis (TBE) in the County of Stockholm. (Open circles) Validated random points from the model.
Host species, TBE, and climate
The number of European hares and roe deer shot varies with years within Stockholm County, but both species have declined continuously from a peak in 1984 for hares and 1992 for roe deer. The present number of roe deer shot is about one-fourth of that in the peak year and for hares one-eighth that of the peak year. During the same time period, red fox has increased from a low in the mid-1980s. The long-term variation in roe deer and European hares is synchronized with the number of foxes and positively correlated with a time lag of 8–15 years, suggesting a long-term periodicity between the species (Fig. 3a, b). The number of European hares is negatively correlated with the number of foxes (log[hare]=32.33−10.29 * log[red fox], F=15.00, p<0.0001) and roe deer are negatively related to number of foxes (log[roe]=19.6−0.71 * log[red fox], F=16.3, p<0.0001). This long-term interaction, over decades, between a predator, the red fox, and its main prey species may have implications for the dynamics of tick infestation measured in number of human TBE cases. A multivariate model with stepwise elimination of variables, using log(TBE) cases as dependent variable, log(number of European hares), log(number of roe deer), log(number of red foxes), summer temperatures, and winter NAO index as independent variables revealed a significant model (F=28.91, p<0.0001, df=2; Fig. 4). The best model explaining number of TBE cases included log(number of hare) and log(number of red foxes), while summer temperature, winter NAO index, and log(number of roe deer) were excluded from the model. The most parsimonious model could be written (log[TBE]=14−0.45 * log[hares] −3.62 * log[red foxes], Akaike information criterion [AIC]=−108.11). The number of European hares and red foxes explained 64.4% of the variation in TBE cases and showed that when numbers of hares and red foxes are both are in low numbers more TBE cases are expected (Fig. 4). The number of European hares is the single most important variable explaining number of TBE cases (61.6%), because red fox as a single variable was not significant and TBE cases are negatively related to number of shot hares (Fig. 5).
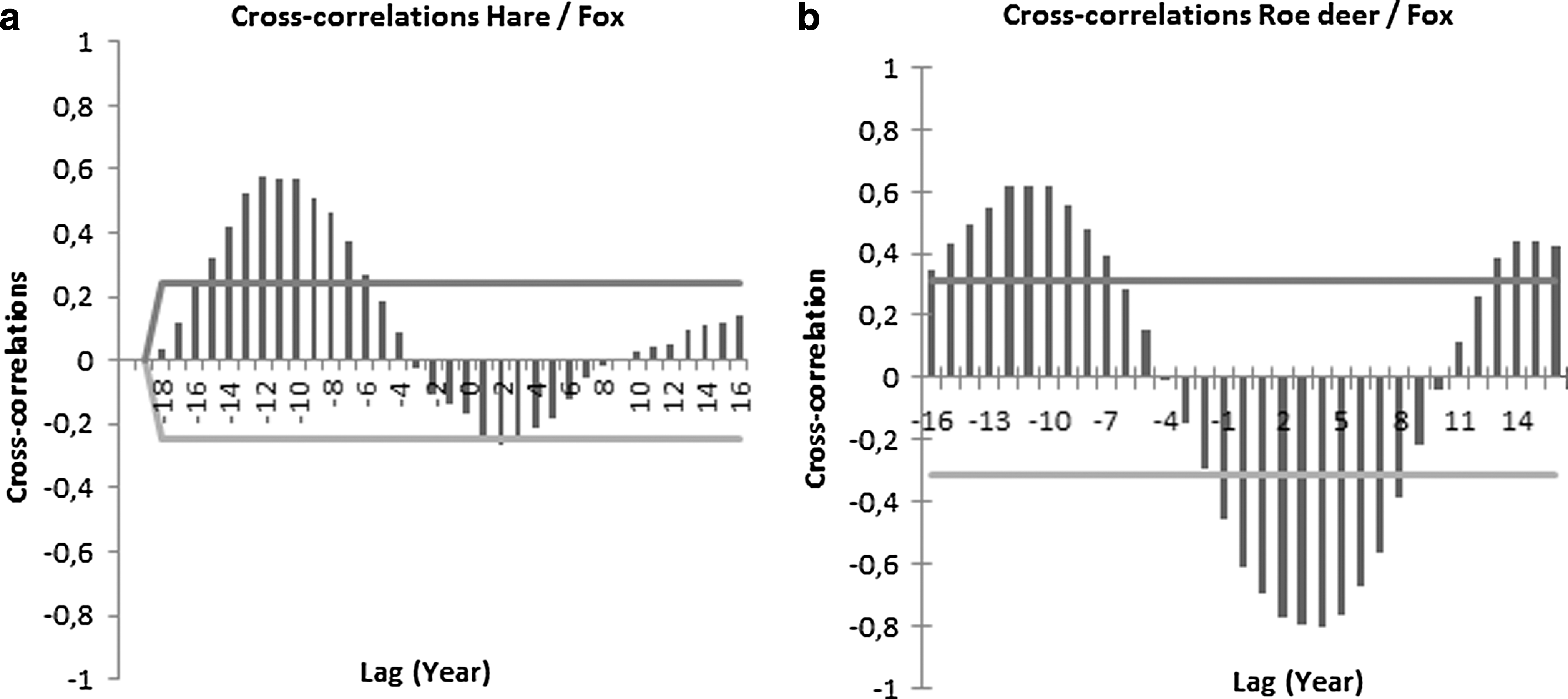
Cross-correlation between number of European hares shot (
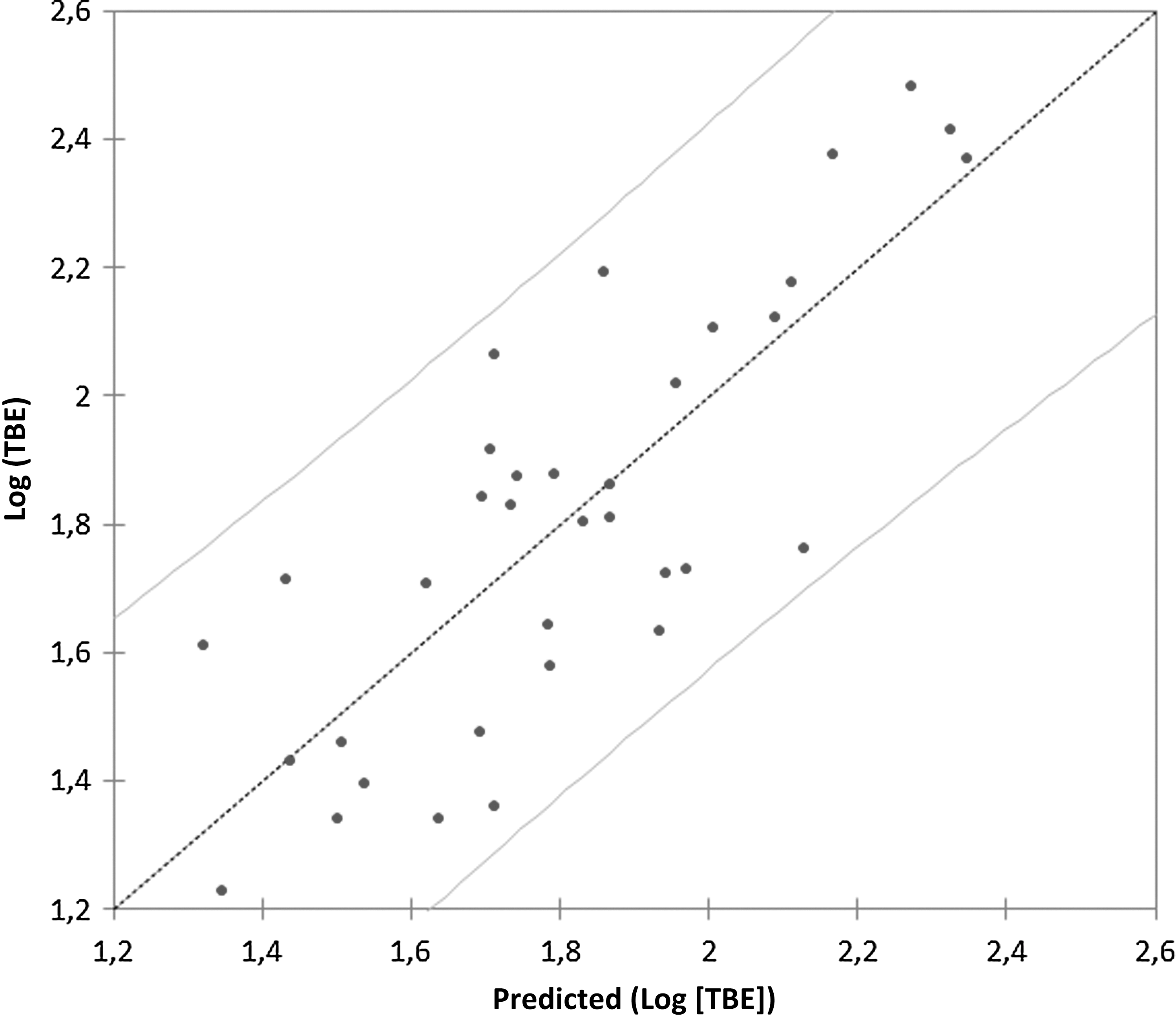
Predicted number of tick-borne encephalitis (TBE) cases from the model with number of European hares and number of red foxes as explanatory variables. Model log(TBE)=14−0.45 * log(hares) −3.62 * log(red foxes), p<0.0001, r 2=0.644.
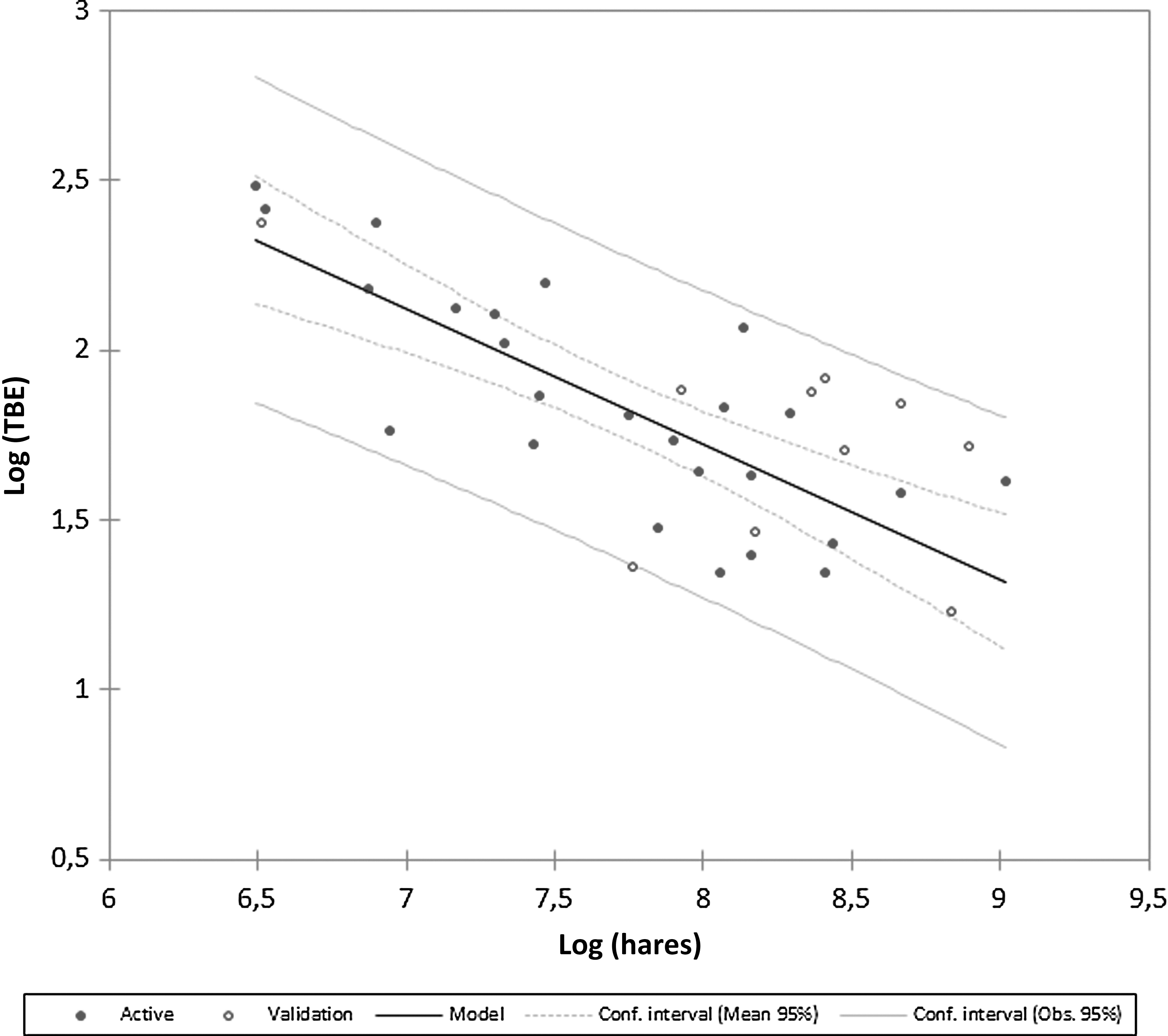
Number of tick-borne encephalitis (TBE) cases in relation the number of European hares in the hunting bag. The model was validated by using random data (open circles).
Discussion
The transmission of TBE to humans from ticks is an increasing threat to human health, and it is important to understand driving forces for disease transmission, especially in view of climate change. The increased number of human cases in Sweden and elsewhere depends on many different factors in the environment, making understanding of disease dynamics a challenge (Jaaskelainen et al. 2010). Among the factors that are expected to be of importance are variations in climate and temperatures because many biological and ecological processes are temperature dependent (Stenseth et al. 2002). Other factors that may influence transmission to humans are human behavior related to both climatic and sociological factors—how people spend their time outdoors and are exposed to infected ticks (Randolph 2010). More importantly, interactions between tick population ecology and host animal populations modulate the dynamics by heterogeneity in transmission between ticks and hosts and effects by predation on hosts (Randolph et al. 2004). The system could not be viewed as in equilibrium or steady state at any point in time. In this study, it was shown that wildlife species that act as hosts for ticks and their population ecology are more important factors determining the number of TBE cases in the County of Stockholm than temperature or large-scale climate index. These results support earlier analyses that climate factors or large scale climatic system were not strong factors to explain variation in TBE cases across Europe (Randolph 2010, Sumilo et al. 2007). This is somewhat surprising because mean summer temperature has increased during the period studied. Even though temperature may be an underlying factor in tick biology, it is masked by other interactions.
The life cycle of ticks is complex because they live both free in the environment and attached to a host. They require several blood meals from several mammalian hosts at different stages in their life cycle to succeed (Nutall et al. 2000). TBE transmission to humans as shown here is more dependent on the number of hosts and effects by predation on host populations than on abiotic factors. This implies that infected tick populations and transfer of virus to humans are a result of an ecological interaction web. In one study on chipmunks (Tamias striatus), it was shown that when the population was low tick larval infestation per host was high (Schmidt et al. 1999). This was also indicated in this study because both hares and roe deer show more TBE cases per individual when populations are low.
The analysis performed here has the drawback that the actual tick population in nature is unknown, and it is not known if the proportion of infected ticks increases in the population or if the prevalence stays the same while the total tick population increases. Irrespective of this fact, the number of human TBE cases in relation to abundance of hosts for ticks speaks in favor for a dilution effect, operating both on diversity and population density (Ostfeldt and Keesing 2000). Abundance of potential hosts is crucial for establishment of infected tick populations, thus it requires knowledge about population dynamics for critical factors in the ecosystem. Many small and medium-sized animals can act as reservoirs for the virus and hosts for the ticks (Gray et al. 2002). It is commonly believed that roe deer and voles are important in this respect, but circumstantial evidence is lacking or importance is inferred from laboratory studies (Gray 1998, Lindgren and Gustafson 2001). Here the analyses of time series show that European hares and red foxes are important actors in the disease dynamics of TBE that have not been previously anticipated. It is commonly believed that a high number of competent hosts facilitate a higher density of ticks and corresponding higher proportion of infected ticks in the population (Randolph et al. 2004, Jaenson et al. 2012). One genus of small rodents, Apodemus, has been shown to be the only competent transmission host, and that co-feeding by larvae and nymphs on animals of this genus is necessary to increase the number of infected ticks (Tälleklint and Jaenson 1993, Randolph et al. 2004, Sumilo et al. 2007). The extent of co-feeding is a density-dependent process, and the likelihood of co-feeding is increasing with few hosts available. An increase in infected tick population requires a low density of host populations but with many ticks per individual host, as has been shown here (Calvet and Leirs 2005). This situation has been formulated as the dilution hypothesis, where high host species diversity and high population numbers dilute the incidence of TBE-infected ticks on any given host at a particular time (Ostfeldt et al. 2006).
Conclusions
From the results presented here, it is clear that summer climate is only a minor factor determining TBE cases in Sweden compared to ecological interactions in the host community, such as predator–prey interactions. A low abundance of hosts and few potential host species increase the risk to humans of TBE infection. It is of interest to collect further information, especially about the population size of infected ticks in relation to uninfected populations, and to include other potential host species in the analysis. The results presented here suggest that TBE transmission is largely determined by interspecific interactions across a tick–prey–predator system.
Footnotes
Acknowledgments
Thanks are extended to Jonas Kindberg at the Swedish Association for Hunting and Wildlife Management, who provided harvest data for the mammalian species, and to Anders Moberg at Stockholm University for the temperature data from the Stockholm Observatory. The Swedish Institute for Disease Prevention provided data on human TBE cases, and climate data was provided by James Hurrell at the National Center for Climate Research, United States (NCAR). Kjell Danell provided valuable comments on the manuscript.