
Abstract
Understanding how human land use patterns influence mosquito ecology and the risk of mosquito-borne pathogens is critical for the development of disease management strategies. We examined how different environments influenced mosquito species composition, abundance, and West Nile virus (WNV) infection rates in central Illinois. Using a combination of gravid traps and CDC light traps, adult mosquitoes were collected every other week from June 24 to September 16, 2012, in four major land use categories—row crops, prairies, forest fragments, and residential neighborhoods. The mosquitoes were identified to species morphologically, and pools of pure and mixed Culex mosquitoes (primarily Culex pipiens and Culex restuans) were tested for WNV-RNA by qRT-PCR. Mosquito species diversity was significantly higher in forest habitats compared to residential, agricultural, and prairie land use categories. All the four landscape types were equally important habitats for WNV vectors Cx. pipiens and Cx. restuans, contrary to previous findings that these species principally inhabit the residential areas. WNV-infected mosquito pools were observed in all land use types, and the infection rates overlapped among land use categories. Although our findings support the importance of residential habitats for WNV transmission to humans, they also establish that prairie, row crops, and wood lots are potentially important refuges for enzootic transmission. This is particularly important in urban ecosystems where these land use categories are small, interspersed fragments serving as potential refuge sites during periods of low rainfall.
Introduction
M
The goal of this study was to examine how land use patterns influence mosquito species diversity and abundance and the risk of WNV transmission in Champaign County, Illinois. WNV was first reported in New York City in 1999 and disseminated rapidly to become the most common mosquito-borne disease in the United States. In 2002, the United States recorded the world's largest outbreak of WNV encephalitis, with Illinois accounting for 21% of the total cases, the most among all states. Geographic variations in human WNV disease incidence and the risk of WNV transmission are well documented, with human land use as one of the risk factors (Yiannakoulias et al. 2006, Bowden et al. 2011, Kilpatrick 2011). However, the impact of human land use on the risk of WNV transmission has not been studied explicitly at fine spatial scales.
Much of the Midwestern United States is dominated by four United States Geological Survey (USGS) land cover categories: (1) Cultivated row crops, characterized by agricultural monocultures such as corn, soybean, and wheat; (2) native and restored prairie, commonly characterized by grasses, sedges, and asters; (3) forest fragments, containing mixed woody deciduous trees (e.g., red oak, Quercus rubra; sugar maple, Acer saccharum), coniferous trees (e.g., white pine, Pinus strobus), and exotic, invasive plants (e.g., bush honeysuckle, Lonicera maackii); and (4) developed residential neighborhoods (Lubowski et al. 2006). The effect of these land covers on mosquito species diversity, abundance, and the risk of WNV transmission are poorly understood, despite their potential to modify availability and abundance of oviposition sites, blood meal hosts, and sugar meal sources for multiple endemic mosquito species. Most of the previous studies have tended to focus exclusively on either Culex pipiens (Diptera: Culicidae) and Cx. restuans and their role in WNV transmission in residential areas (Gu et al. 2006, Kunkel et al. 2006) or on nuisance and medically important Aedes mosquitoes including Aedes vexans, a common floodwater nuisance mosquito (Horsfall et al. 1975), Ae. triseriatus, the vector of La Crosse virus (LACV) (Walker et al. 1997), and two recently introduced mosquito species, Ae. albopictus and Ae. japonicus, that have the potential to negatively impact the distribution and abundance of native mosquito species (O'Meara et al. 1995, Armistead et al. 2008, Bevins 2008) and to vector a suite of native and exotic arboviruses including WNV, dengue, La Crosse encephalitis virus (LACV), eastern equine encephalitis, and Japanese encephalitis (Rosen et al. 1985, Mitchell 1995, Sardelis and Turell 2001, Sardelis et al. 2002a, Sardelis et al. 2002b, Turell et al. 2005).
Because human land use patterns can potentially alter the ecology of mosquito vectors and the risk of arbovirus transmission (Brown et al. 2008, Savage et al. 2008, Kilpatrick et al. 2011, Crowder et al. 2013), a proper understanding of how the four common land use categories in the Midwestern United States influence mosquito ecology and the risk of WNV transmission is essential. This study addressed two related questions regarding the effect of land use on mosquito biodiversity and the risk of WNV transmission in Champaign County, Illinois. First, how does mosquito species diversity and abundance compare among land use categories? Second, is the risk of WNV transmission similar across land use categories? The findings of this study may allow us to predict and respond to land use categories that pose particularly high risk for transmission of WNV and other mosquito-borne pathogens, including the long-term implementation of an Integrated Vector Management (IVM) program locally and in comparable mixed landscapes throughout the Midwestern United States.
Materials and Methods
Adult mosquitoes were collected in Champaign County, Illinois, an urban ecosystem with a human population density of 74 people/km2. Arboviruses detected in Illinois and their vectors include WNV, primarily vectored by Cx. pipiens and Cx. restuans (O'Leary et al. 2004); LACV, vectored by Ae. triseriatus (Mitchell et al. 1998); eastern equine encephalitis, vectored by Culiseta melanura (Scott et al. 1984); and St. Louis encephalitis (SLEV), vectored by Cx. pipiens and Cx. restuans (Creech 1977). Like many cities in the Midwest, Champaign is characterized by a mixed landscape with four general land use categories—cultivated row crops (particularly corn and soybean), restored prairies, forest fragments, and residential neighborhoods—within a narrow spatial radius (the Euclidean distance between our most remote study sites was ∼16 km). Study sites were established in ecosystems representing these habitat types, with three sites each in agricultural, prairie, forest fragment, and residential land use categories (Fig. 1).
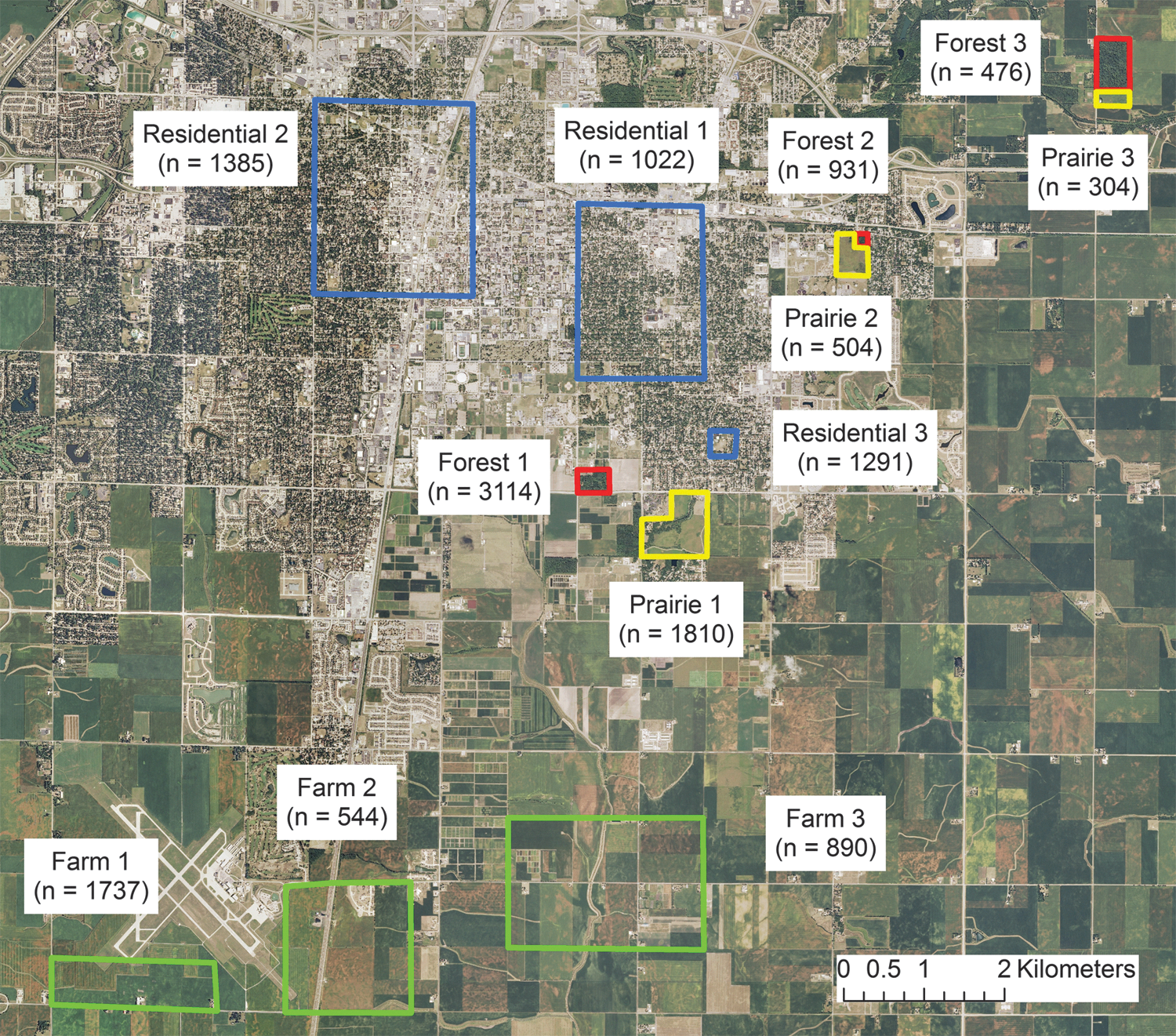
Locations of study sites in Champaign County, Illinois, and number of adult female mosquitoes collected per site. Land use categories represented were agricultural, forest fragment, prairie, and residential. Color images available online at
Three pairs of grass infusion-baited gravid traps and CDC light traps not baited with CO2 were operated in each of the three study sites of a landscape. The two trap types were placed approximately 10 meters apart, and the distance to the next pair of traps was at least 100 meters, with CDC light traps at 1.5 meters above the ground. CDC light traps are designed to collect a representative cross-section of the mosquito community, including males and females of mixed parity (McCardle et al. 2004), whereas gravid traps are designed to primarily collect gravid Cx. pipiens and Cx. restuans females seeking stagnant water sources to oviposit. There also is an inherent difference in the likelihood of detecting WNV in these traps, because CDC light traps capture many nullipars whereas most mosquitoes captured in gravid traps have taken at least one blood meal. Mosquito collections were conducted once every 2 weeks between June 24 and September 16, 2012. The average trap night lasted from 1600 to 0900 h. Adults were identified morphologically to species on a chill table (Darsie and Ward 2005). Adult females of each species were pooled by site and date of collection in groups of up to 50 individuals and tested for WNV viral RNA by a Taqman RT-PCR assay (Lanciotti et al. 2000, Lampman et al. 2006).
Data were analyzed using R version 2.12.1 (R Foundation for Statistical Computing, Vienna, Austria). Data were checked for normality and homogeneity of variance, and abundance data was log+1-transformed to meet these assumptions. Female mosquito species diversity among different land use categories was quantified using the Shannon diversity index (H), a community diversity measure that accounts for both the abundance and the evenness of the species present in a given locale. Spatial variation in mosquito abundance between different habitats was assessed using mixed linear models with repeated measures. To differentiate between net mosquito abundance and abundance of medically important species, two models were generated using cumulative mosquito abundance and Cx. pipiens and Cx. restuans abundance as the responses.
Maximum likelihood estimates for infection rates were calculated using the Excel (Microsoft, Redmond, WA) add-in for Pooled Infection Rate version 3.0 (Biggerstaff 2003). The data included the combined WNV results for female pools of Cx. pipiens, Cx. restuans, and mixtures of the two Culex species. To evaluate differences in transmission between categories, the infection rate estimate was multiplied by the total number of mosquitoes to provide the best estimate of number of infected mosquitoes at each site and time period. A multiple proportion chi-square analysis was performed to evaluate spatial and temporal differences in the proportion of infected to uninfected mosquitoes. The estimated number of infected mosquitoes was standardized by the number of trap nights per location, category, or week and is presented with infection rate (and 95% confidence limits).
Results
Mosquito species composition, abundance, and diversity
Overall, a total of 14,007 adult female mosquitoes were collected at the 12 study sites during the 7-week period (Table 1). The largest number of mosquitoes was collected in forest fragments (n=4561), followed by residential habitats (n=3701), agricultural habitats (n=3138), and tallgrass prairie habitats (n=2607). From this total, 5121 Culex specimens could not be identified to species due to loss of morphological characteristics during collection. The remaining specimens yielded 18 mosquito species comprising six species of Culex, five species of Aedes, three species of Anopheles, and one species each of Coquillettidia, Culiseta, Orthopodomyia, and Uranotaenia (Table 1). Seventeen of the 18 mosquito species were collected in forest sites compared to 11, 10, and 9 species, respectively, in prairie, residential, and agricultural sites. Although the number per species varied between trap types, the total number of mosquito species collected in CDC light traps and gravid traps was identical across the four study sites (8 vs. 9, 10 vs. 10, 15 vs. 14, and 10 vs. 10 species for agricultural, prairie, forest, and residential sites, respectively, in CDC and gravid traps).
Values in parentheses indicate percentage of column total.
Cx. pipiens was the most dominant species in all land use types and was mainly collected in gravid traps. There were no significant differences in abundance of this species among land use types (F=2.20; degrees of freedom [df]=3, 8; p=0.09). In all study sites, it was common to collect some mosquito species in one trap type but not the other. In agricultural sites, for example, Cx. salinarius was only collected in gravid traps whereas Ae. vexans and Culiseta inornata were collected only in CDC light traps. Similarly in forest sites, Cx. erraticus, Cx. territans, Anopheles walkeri, and Uranotaenia sapphirina were collected in CDC light traps, whereas Cx. salinarius and Ae. albopictus were collected in gravid traps. Although there were no significant differences in Shannon's species evenness among land use types, Shannon's species diversity index was significantly higher in forest sites compared to the other land use categories in CDC light trap collections (F=10.85; df=3, 32; p<0.01, Table 2). However, there was considerable variation between collection sites in some land use categories.
Values in parentheses indicate species richness.
Within-land use variations in mosquito abundance
We observed statistically significant within-habitat variation among different sites in the same land use category. Within the agricultural land use category, total mosquito abundance (F=12.21; df=2; p<0.01) and Culex spp. abundance (F=12.18; df=2; p<0.01) were significantly higher at Farm 1 than at the Farm 2 and Farm 3 sites (Fig. 1). Within the prairie land use category, total mosquito abundance (F=13.25; df=2; p<0.01) and Culex spp. abundance (F=13.87; df=2; p<0.01) were significantly higher at Prairie 1 than at the Prairie 2 and Prairie 3 sites. Within the forest fragment land use category, total mosquito abundance (F=16.16; df=2; p<0.01) and Culex spp. abundance (F=18.84; df=2; p<0.01) were significantly higher at Forest 1 than at the Forest 2 and Forest 3 sites. No within-habitat variation was observed among the sites in the residential land use category (F=0.29; df=2; p=0.75), suggesting a broad uniformity in Culex abundance between urban sites.
Land use and WNV
The infection rates in the Cx. pipiens and Cx. restuans samples varied by land use category, with 26.5 infected Culex per 1000 in forest fragment, 21.8 in residential, 16.6 in agricultural, and 14.9 in prairie habitats (Table 3). The proportion of infected mosquitoes in forest fragments was significantly greater than in the agricultural and prairie landscape categories, but it was not different from the residential category (χ2=14.91; df=3; p<0.01). The infection rate estimates and estimated number of infected Culex at each site yielded different patterns of risk (Table 3). Infection rates did not compensate for differences in mosquito abundance; thus, some sites with low infection rates (i.e., Prairie 1 site) actually had a higher number of infected mosquitoes despite the lower rate. Each of the land use categories had one site with a higher number of infected mosquitoes that would have been masked by considering infection rates only (Table 3). The number of infected Culex per trap was not significantly different among residential, agricultural, prairie, and forest sites; however, Forest 1 had a four-fold or greater number of infected Culex (4.05 per trap night) than the other sites (Table 3). If this outlier site was removed, a one-way analysis of variance (ANOVA) found significant differences (F=15.5; df=3, 7; p<0.01) with residential having 1.19 infected Culex per trap night, which was significantly more than forest fragment (0.53) and prairie (0.50), but not different from infected Culex in the agricultural land use category (0.88).
Letters represent differences between proportion of infected and uninfected mosquitoes in each land use category by multiple chi-squared analysis.
WNV, West Nile virus; MLE-IR, maximum likelihood estimation infection rates.
The pattern of peak transmission varied between infection rate estimates and number of infected mosquitoes per week. Temporally, there was a peak in infection rates the week of August 6; however, the proportion of positive mosquitoes was equivalent in the weeks of July 23 and August 6, but significantly different from the previous two weeks (Fig. 2). In contrast, the number of positive mosquitoes per trap night peaked at the week of August 6 and declined thereafter, despite an apparent infection rate rebound in the week of September 17.
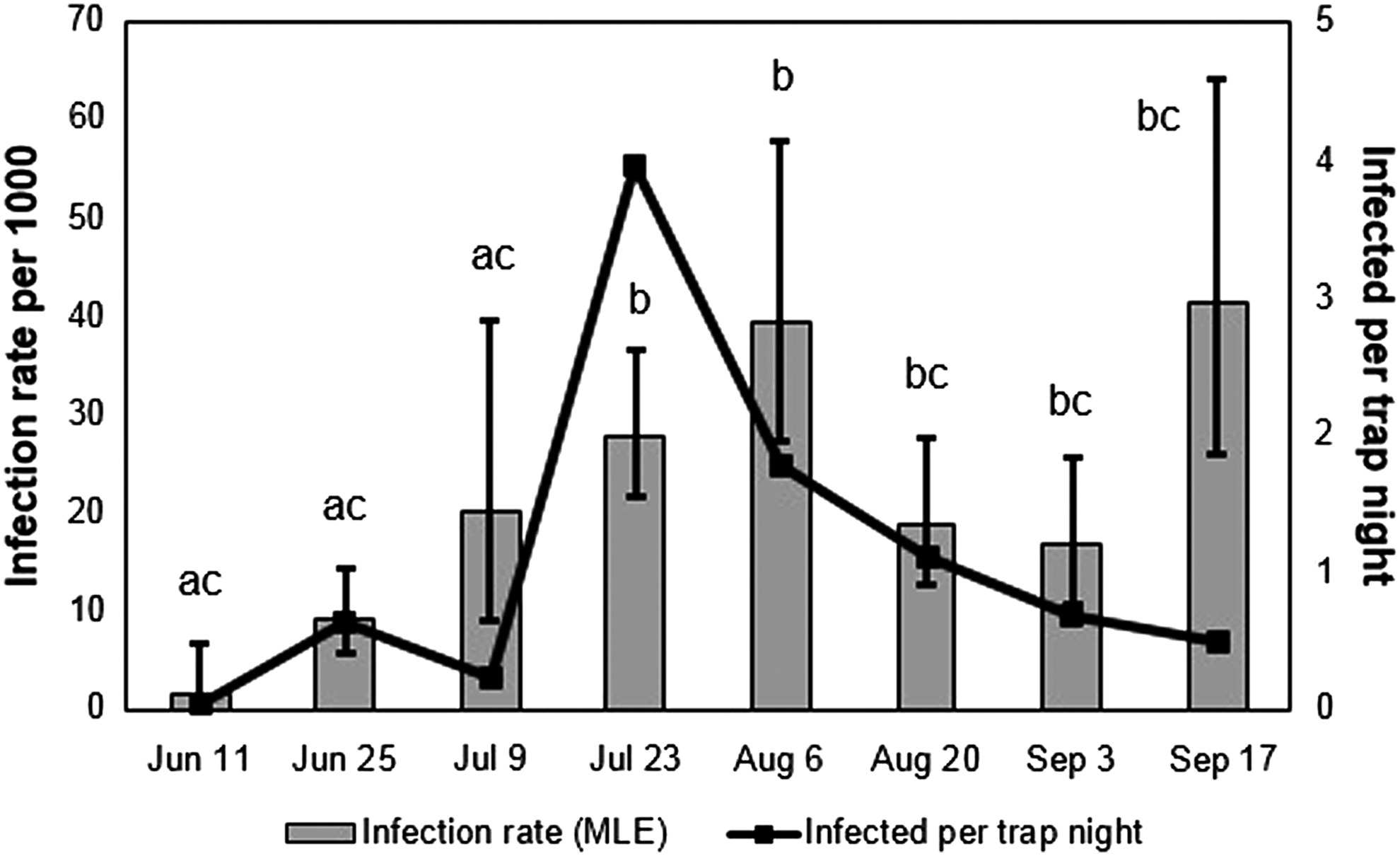
Temporal variation in infection rates (bars) and number of infected Culex pipiens, Cx. restuans, and unidentified Culex spp. (line) for all sites and categories combined. Letters represent differences in the proportion of infected to uninfected mosquitoes. MLE, maximum likelihood estimation.
Discussion
The results support the hypothesis that land use patterns in a moderate-sized urban area (Urbana-Champaign) of central Illinois do affect species diversity and abundance in different land use categories. Species diversity was significantly higher in forest habitats compared to residential, agricultural, and prairie land use categories in both CDC light trap and infusion-baited gravid trap collections. We did not quantify the type and abundance of larval habitats in each landscape type; however, previous studies suggest that forest landscapes may encompass a greater diversity of larval habitats (e.g., tree holes, ditches, vernal pools, leaf axils), favoring higher mosquito species diversity, than urban landscapes, which are often associated with a higher abundance of vector species, especially Culex and Ochlerotatus spp. associated with artificial containers (Keating et al. 2004). Furthermore, forest fragments may provide a greater diversity of blood and sugar sources for adult mosquitoes compared to other habitats. For example, Illinois urban residential locales and agricultural monocultures tend to be dominated by only four avian species, the American robin (Turdus migratorius), house sparrow (Passer domesticus), mourning dove (Zenaida macroura), and northern cardinal (Cardinalis cardinalis), whereas avian diversity is higher in wooded sites (Hamer et al. 2008, Loss et al. 2009). The role of host diversity in modifying arbovirus transmission is mixed. In northern Illinois, no relationship was found between WNV prevalence and avian diversity (Loss et al. 2009), whereas other studies at different spatial scales suggest that an increase in blood host diversity may lower transmission of WNV (reviewed in Ostfeld and Keesing 2012). We lacked the host diversity by land use information to explore this relationship.
Climatic conditions directly influence the abundance of larval habitats for floodwater and container-breeding mosquitoes (Patz and Reisen 2001). According to the Illinois State Water Survey, Champaign County recorded unusually high temperatures and low precipitation in 2012, particularly in July and August. The summer of 2012 was a drought season in Illinois, with a maximum rainfall accumulation of 2.78 cm over 24 h and a cumulative rainfall of 13.22 cm during the study period compared to a 20-year average cumulative rainfall of 32.94 cm over the same time frame (Illinois State Water Survey 2013). The lack of rainfall during these months probably biased species distributions, and the reduction in overall larval habitats may be partially responsible for the presence and dominance of Cx. pipiens across all land use categories. However, our results reflect those that would be considered favorable for WNV outbreaks, namely above-average temperatures and below-average precipitation (Epstein et al. 2001, Ruiz et al. 2010).
Broadly posed, all four landscape categories were equally important habitats for the primary northeastern WNV vectors Cx. pipiens and Cx. restuans. Considerable variations in mosquito abundance were observed within the forest, prairie, and agricultural landscape categories, suggesting that mosquito populations in these habitats may be influenced by the surrounding environment. Farm 1, which was located proximate to Willard airport and the University of Illinois golf course, and Forest 1 and Prairie 1, which were located within urban ecosystems, had higher mosquito abundance than the remaining sites in their respective land use category. This observation confirms previous findings that Cx. pipiens and Cx. restuans tend to be more abundant in human-dominated environments (Andreadis et al. 2001, Fonseca et al. 2004), yet they are also broadly distributed in a variety of landscapes. These findings indicate that the common approach of grouping transmission data over broad spatial scales, like counties, may not be appropriate, and that examining smaller intraurban habitats is key to identifying important co-variates in WNV transmission. Our results suggest that all of the land use patterns may be refuges for Culex mosquitoes during drought-like conditions. The importance of rainfall in the determination of larval mosquito production and adult distribution is well established in a variety of ecosystems (Koenraadt and Harrington 2008). Rainfall may impact mosquitoes positively by creating new habitats or by extending the duration in which they remain available for immature development, and rainfall can interact negatively by flushing larvae out of their larval habitats.
The data yielded mixed support for the second main hypothesis that arbovirus transmission is affected by land use category. WNV-infected mosquito pools were observed in all land use types, and the infection rates overlapped among land use categories. However, the number of WNV-infected mosquitoes per trap site was two- to four-fold higher at the University of Illinois South Farms wood lot (Forest 1) compared to the other sites. This was atypical and may reflect the closeness of this site to residential areas, a dairy cattle farm, and a local abundance of artificial containers in the area (Morris et al. 2007). Once this outlier was excluded from the analysis, the residential sites had the highest number of infected mosquitoes, although not significantly different from the agricultural land use category. This pattern of more infected mosquitoes (hence increased risk) tends to agree with the urban and rural clustering patterns of human and horse cases (Ward et al. 2005). We compared infected to uninfected mosquitoes collected at each site rather than just infection rates (the proportion of positive mosquitoes), because the spatial and temporal variation in abundance of mosquitoes is also a key risk factor in arbovirus transmission (Kilpatrick et al. 2005, Nasci et al. 2005). The difference between infection rate and number of infected mosquitoes was particularly evident in temporal comparisons (Fig. 1), which showed a peak infection rate in the week of August 6 and a peak in number of infected mosquitoes in the week of July 23 for all sites combined. The abundance of infected vectors in all land use categories further demonstrates the potential for various areas to act as harborage of enzootic transmission and that fine-scale structure (Forest 1) may have a significant impact on arbovirus transmission. These results suggest that under drought-like conditions, WNV vector and transmission surveillance may need to encompass a variety of land use categories.
In summary, we observed significantly higher mosquito species diversity in forest landscapes compared to other land use categories. We also found that all four land use types examined in this study may present similar risks for WNV transmission during drought years and that risk may vary within a category. These results may be directly applicable to mosquito management practices in central Illinois and comparable heterogeneous landscapes, where much of vector control focuses intensively on human-inhabited areas and little emphasis is placed on other land use sectors within the broader urban ecosystem. Future research in these environments may address questions, including whether mosquito blood meal host diversity varies similarly with land use history and structure and whether the results of this study can be generalized to regions under different weather and climate conditions.
Footnotes
Acknowledgments
We thank Nina Krasavin, Millon Blackshear, Allison Montgomery, Rosa Alfaro, Macey Brusich, James Ricci, Dan Bernard, Thorsten Hansen, Travis Weinsheim, and Andrew Donelson for their technical assistance. This study was supported by the Used Tire Fund and Emergency Public Health Fund from the State of Illinois.
Author Disclosure Statement
No competing financial interests exist.