
Abstract
Two rickettsial isolates, HM-1 and HM-2, were isolated from Haemaphysalis megaspinosa collected in Japan in 2006 and 2011, respectively. The isolates were analyzed by DNA sequences of the outer membrane protein A gene, the outer membrane protein B gene, the citrate synthase gene, the genus Rickettsia-specific outer membrane protein 17-kDa gene, the 16S ribosome RNA gene, and the PS120 protein gene (“geneD”). HM-1 was identified as Rickettsia tamurae. HM-2 matched most closely with ‘Candidatus Rickettsia kotlanii’ DNA, which has only been reported from H. concinna in Hungary. This is the first report of isolation in Japan of the agent genetically similar to ‘Candidatus R. kotlanii,’ which belongs phylogenetically to the spotted fever group Rickettsia. Our study shows the possibility that ‘Candidatus R. kotlanii’ can be carried by at least two tick species. Furthermore, because the Rickettsia sp. has been found two distant countries, Hungary and Japan, it has potential for wider distribution.
Introduction
R
Rickettsia tamurae was first isolated from Amblyomma testudinarium in Japan. It belongs to SFG phylogenetically and was recently associated with lesional skin inflammation in an afebrile human (Imaoka et al. 2011). R. tamurae is more prevalent in Japan than R. japonica (Gaowa et al. 2013).
‘Candidatus Rickettsia kotlanii,’ a new SFG Rickettsia has been discovered in Haemaphysalis concinna in Hungary (Sreter-Lancz et al. 2006, Hornok et al. 2010). However, isolation of the microbe has not yet been reported.
Haemaphysalis megaspinosa is distributed throughout Japan and appears to have an affinity for humans, on the basis of a report of a tick bite case (Seishima et al. 2000). In Japan, H. megaspinosa is known to carry Rickettsia, as shown by detection of R. japonica (Katayama et al. 2001) and Rickettsia sp. Hf332 (closely related to R. japonica) DNA (Ishikura et al. 2003). Therefore, H. megaspinosa is likely a vector of other Rickettsiae as well. Here we report isolation of two new rickettsial isolates from H. megaspinosa collected in Japan.
Materials and Methods
Ticks were collected at Nakadori and Nakae islands of the Goto archipelago, Nagasaki prefecture in April, 2006, and at Shin-hidaka, Hokkaido in May, 2011, as part of routine surveillance in Japan. Ticks were identified morphologically to species and stage (Yamauti et al. 1971), and H. megaspinosa were used for further investigation.
Rickettsia isolation was performed by the shell vial method. Ticks were homogenized individually in sucrose–phosphate–glutamate buffer and inoculated into L929 cells. The inoculated cells were cultured until Rickettsia-like particles could be confirmed using phase-contrast microscopy (magnification, 400×). Immunoperoxidase staining was also performed to detect Rickettsia particles in the cells using SFG-specific monoclonal antibodies S3 and X1 (Oikawa et al, 1993).
DNA was extracted from Rickettsia-infected cells, primary culture at 7 days after inoculation of the samples, using the Puregene DNA extraction kit (Qiagen, Valencia, CA). For nested PCR, primer pairs R1/R2 and Rr17.61p/Rr17.492n (Sakamoto et al. 2013) were used for the genus Rickettsia-specific outer membrane protein gene (17-kDa); Rr190.70p/pre-amp1 (CCTCCATCTATATATTTCCTGCAAGTG) and Rr190.70p/Rr190.602n (Regnery et al. 1991) were used for the outer membrane protein A gene (ompA); and Cs2d/CsEndr and RpCS.877p/RpCS.1258n (Sakamoto et al. 2013) were used for the citrate synthase gene (gltA). Primer pair 120-M59′/120-807′ was used for rOmpB protein encoding gene (ompB) (De Sousa et al. 2006) and primer pairs D1f/D928r, D767f/D1390r, D1219f/D1876r, D1738f/D2482r, and D2338f/D3069r were used for PS120 protein gene (“gene D”) (Sekeyova et al. 2001). The TaKaRa Bacterial 16S rDNA PCR kit (TaKaRa, Shiga, Japan) was used for 16S ribosome RNA gene (rrs) detection. PCR was performed using conditions described in the original articles.
Direct sequencing was performed using a BigDye Terminator v3.1 cycle sequencing kit (Applied Biosystems, CA) on an ABI Prism 3130 Genetic Analyzer (Applied Biosystems, CA). Sequenced data were aligned using ATCG software (Genetix) and identified by comparison with GenBank using the BLAST program (
Results
A total of 53 H. megaspinosa (eight larvas, 33 nymphs, one female, and 11 males) were collected from the Goto archipelago. Of those, 41 ticks (one larva, 31 nymphs, and nine males) were used for isolation, and one isolate, HM-1, was isolated from a male tick from Nakadori Island. At Shin-hidaka, 55 H. megaspinosa (five nymphs, 28 females, and 22 males) were collected; 36 ticks (five nymphs, 18 females, and 13 males) were used for isolation, and one isolate, HM-2, was isolated from a nymph tick. Isolation of the agents was confirmed by 7 days after inoculation of the tick samples to the cells (Fig. 1). The cultures were continued for several passages after confirming the isolations until the isolates were established in vitro. Frozen stocks of the isolates were made for future studies.
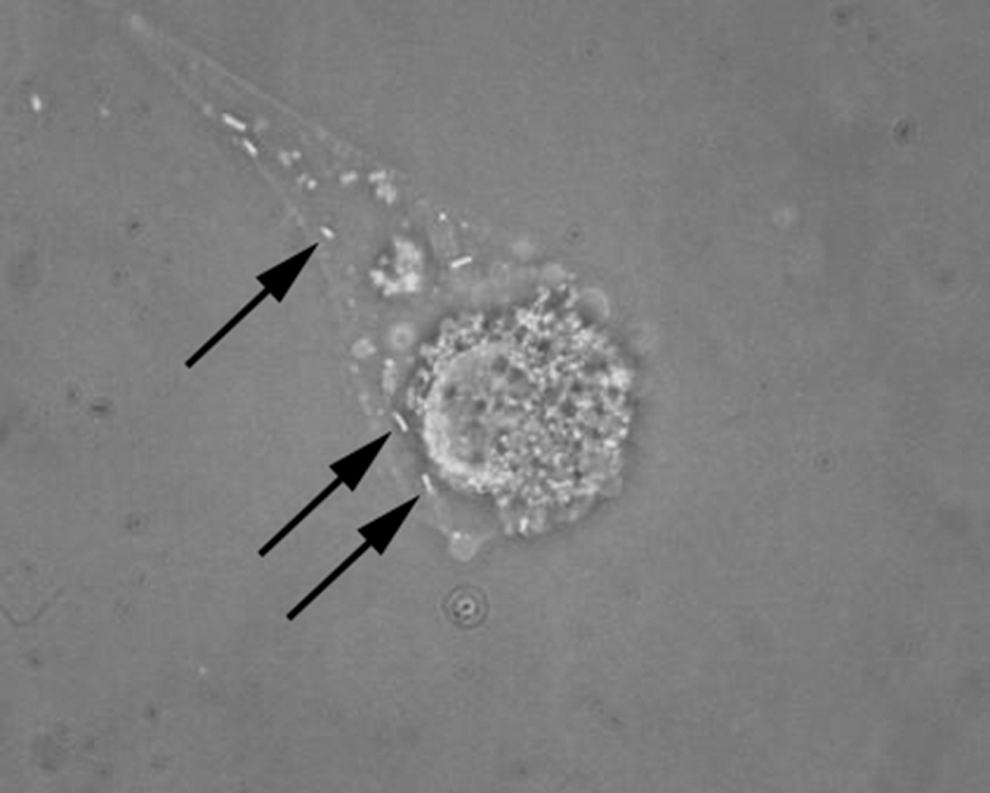
Phase-contrast image of L929 cells infected with HM-2 isolate. Arrows indicate Rickettsial particles. Original magnification, 400×.
HM-1 was identified as R. tamurae from the sequence similarity of ompA (AB812552) (488/488 bp), gltA (AB812551) (1231/1232 bp), and 17kDa (AB812550) (393/394 bp) genes with R. tamurae AT-1 strain (AB114823, AF394896, and AB114825, respectively).
HM-2 was revealed to be genetically similar to ‘Candidatus R. kotlanii’ by the sequence similarities of ompA, gltA, and 16S rRNA genes. The similarity of the ompA gene of HM-2 (AB781706) and ‘Candidatus R. kotlanii’ isolate HcH1 (DQ243938) was 98% (479/489 bp). The gltA gene of HM-2 (AB781705) was 100% (341/341 bp) identical to ‘Candidatus R. kotlanii’ isolate Hu6-2007 (EU853835) and isolate HcH1 (DQ243937). The rrs gene of HM-2 (AB781707) and isolate HcH1 (DQ243936) were 100% (748/748 bp) identical. The 17-kDa gene (AB781704), the ompB gene (KJ619626), and the ‘gene D’ (KJ619625) of HM-2 had no 100% similar gene in the database. The information of the17-kDa gene, the ompB gene, and the ‘gene D’ of ‘Candidatus R. kotlanii’ were not available. The phylogenetic trees are shown in Figure 2.
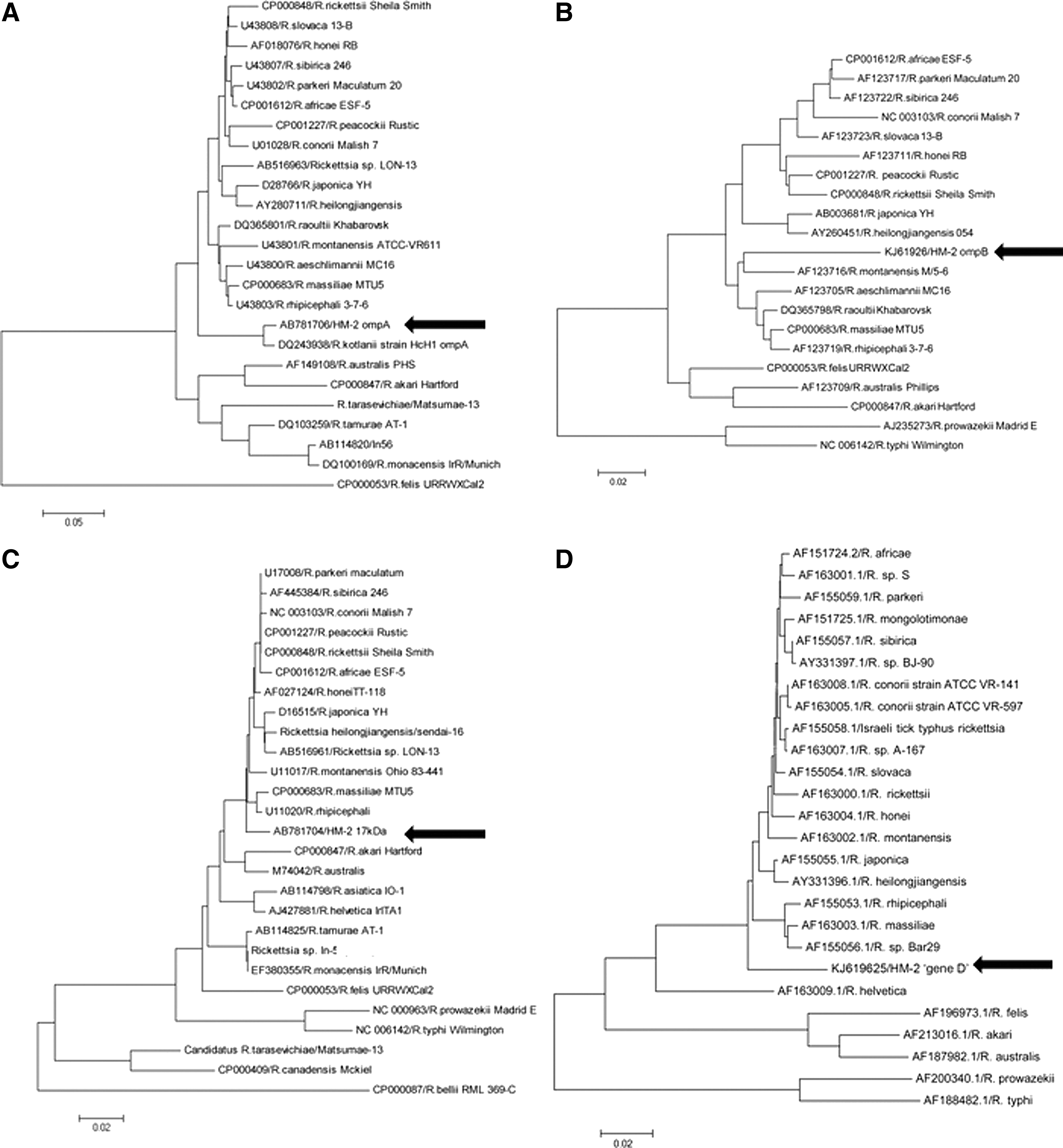
Phylogenetic tree of ompA (
Discussion
We isolated two rickettsial agents from H. megaspinosa collected in Japan. Isolate HM-1 was R. tamurae, which has been commonly isolated from A. testudinarium. Another isolate, HM-2, was genetically similar to ‘Candidatus R. kotlanii’, which has never been reported in Japan. The isolation of HM-2 in Japan indicates the possibility of broad geographical distribution of this Rickettsia sp.
‘Candidatus R. kotlanii’ has been described in two publications, and both of them were reported to be from ticks collected in Hungary. The first report demonstrated detection of DNA of a new SFG Rickettsia from two pools of 100 engorged H. concinna pooled samples (Sreter-Lancz et al. 2006). The second report demonstrated detection of DNA from H. concinna collected from vegetation with just one positive in 52 pooled samples (Hornok et al. 2010). The prevalence of a Rickettsia sp. that is genetically similar to ‘Candidatus R. kotlanii’ was 2.8% (1/36) in the present study. However, we speculate that the general prevalence is even lower because we have never previously detected DNA from or isolated this Rickettsia sp. in Japan despite numerous epidemiological studies using ticks (Gaowa et al. 2013). Although the prevalence is low, our study shows the diversity of Rickettsia spp. in Japan. Isolation of the HM-2 strain provides significant public health information: that ‘Candidatus R. kotlanii’ or a similar agent is more widespread than previously thought—not just in Hungary—and that the Rickettsia sp. has at least two host tick species.
The host ticks of ‘Candidatus R. kotlanii’, H. megaspinosa, and H. concinna, both show affinity for humans. This suggests that humans may be at risk of infection. The isolation of the new Rickettsia sp. enables study of its biological characters, including pathogenicity. Because Rickettsia sp. HM-2 and ‘Candidatus R. kotlanii’ belong to the SFG phylogenetically, it is important to know its virulence against humans and/or animals in case this species causes disease and diagnostic tools and therapies are needed.
Footnotes
Acknowledgments
This work was supported by grants of the Japanese Ministry of Health, Labour and Welfare (H21-Shinkou-Ippan-006 and H24-Shinkou-Ippan-008). We are grateful to Dr. Kyle Rueben Taylor for critical reading and editing of the manuscript.
Author Disclosure Statement
No competing financial interests exist.