
Abstract
Escherichia coli strains such as Shiga toxin–producing E. coli (STEC), enteropathogenic E. coli, enterotoxigenic, attaching, and effacing E. coli, and enteroinvasive E. coli cause diarrhea in humans. Although other serotypes exist, the most commonly reported STEC in outbreaks is O157:H7. A cross-sectional study was conducted to isolate and characterize non-sorbitol–fermenting (NSF) E. coli O157:H7 from urban and periurban livestock settings of Morogoro, Tanzania. Human stool, cattle feces, and soil and water samples were collected. Observations and questionnaire interview studies were used to gather information about cattle and manure management practices in the study area. E. coli were isolated on sorbitol MacConkey agar and characterized by conventional biochemical tests. Out of 1049 samples, 143 (13.7%) yielded NSF E. coli. Serological and antimicrobial tests and molecular typing were performed to NSF E. coli isolates. These procedures detected 10 (7%) pathogenic E. coli including STEC (n=7), enteropathogenic E. coli (EPEC) (n=2), and attaching and effacing E. coli (A/EEC) (n=1) strains. The STEC strains had the ability to produce VT1 and different VT2 toxin subtypes that caused cytopathic effects on Vero cells. The prevalence of STEC in cattle was 1.6%, out of which 0.9% was serotype O157:H7 and the overall prevalence of diarrheagenic E. coli in cattle was 2.2%. The serotypes O157:H7, O142:H34, O113:H21, O+:H−, O+:H16, and O25:H4 were identified. One ESBL-producing isolate showed the MLST type ST131. To our knowledge, this is the first finding in Tanzania of this recently emerged worldwide pandemic clonal group, causing widespread antimicrobial-resistant infections, and adds knowledge of the geographical distribution of ST131. Cattle manure was indiscriminately deposited within residential areas, and there was direct contact between humans and cattle feces during manure handling. Cattle and manure management practices expose humans, animals, and the environment to pathogenic E. coli and other manure-borne pathogens. Therefore, there is a need to improve manure management practices in urban and periurban areas to prevent pathogen spread and associated human health risks.
Introduction
D
Once referred to as a rare serotype (Riley et al. 1983), non-sorbitol–fermenting (NSF) STEC O157:H7 is now a public health problem worldwide (Caprioli et al. 2005). STEC O157:H7 typically causes hemorrhagic colitis characterized by severe abdominal pain, watery diarrhea preceding bloody diarrhea, and little or no fever (Germani et al. 1997). About 2–15% of STEC O157:H7 infections progress to HUS characterized by hemolytic anemia, thrombocytopenia, and renal failure (Fitzpatrick 1999, Dundas et al. 2001). Both sporadic cases and epidemic diseases associated with STEC O157:H7 have been reported (Sang et al. 1996, Crump et al. 2002).
Hosts and sources of STEC O157:H7 are diverse and include cattle, sheep, goats, pigs, water buffalo, deer, horse, dogs, rabbits, gulls, pigeons, chicken, meat, milk, poultry and fish products, vegetables, and water (Caprioli et al. 2005, Ferens and Hovde 2011). Cattle are the main reservoir (Chase-Topping et al. 2008) and have been associated with several cases of human infection (Aspán and Eriksson 2012). Adult cattle are tolerant to infection due to lack of vascular receptors for the Shiga toxins produced by STEC O157:H7 (Pruimboom-Brees et al. 2000), whereas ileocolitis in calves following experimental infection is fatal (Dean-Nystrom et al. 1998). Transmission of STEC O157:H7 to humans occurs through the fecal–oral route following ingestion of contaminated food or water (Tuttle et al. 1999, Olsen et al. 2002) or direct contact with contaminated surfaces or animals (Crump et al. 2002).
Urban areas are city areas characterized by a dense human population of mixed age, sex, family and household structure, ethnic, cultural, religious diversity, educational and income levels, and high built-up area with technological and economic advancement. Between the urban and the rural lies a zone of gradual transition from urban to rural, called the periurban area (Simon et al. 2004). Urban and periurban livestock farming is expanding in developing countries primarily due to high demand for animal protein to serve rapidly growing urban populations, but also to generate household income (Mlozi 1997). In Tanzania, there is an increase in both the number of cattle and households engaged in cattle keeping, with such households located in close proximity to households not keeping cattle. Thus, cattle and other livestock manure is deposited indiscriminately in and on the fringes of cities and contaminates the external environment, e.g., cattle defecate when taken around during the day for feeding or when manure is poorly managed at the household level (Lupindu et al. 2012). However, in contrast to well-known health risks associated with poor management of human excreta, little is known about health risks associated with livestock keeping and manure management in urban areas.
The present study, therefore, aimed to estimate the prevalence of NSF STEC O157:H7and other NSF E. coli in cattle, humans, and the associated environment of the municipality of Morogoro, Tanzania.
Materials and Methods
Selection of study subjects and sample collection
The study included smallholder cattle keeping and non-cattle–keeping households of urban and periurban areas of Morogoro municipality, Tanzania. The urban area of Morogoro includes densely populated residential, commercial, industrial, and administrative areas where cattle are also kept. The periurban area lies adjacent to and separates the urban area of Morogoro from the rural settings. It is characterized by dense human population, small cattle herds, and limited crop growing on the urban interface and gradually changes into sparse population, big cattle herds, extensive crop fields, and bushes toward the rural boarder. There is no clearcut demarcation between urban and periurban settings. A herd of cattle belonging to a household was regarded as a cluster. To establish the prevalence of NSF STEC O157:H7, the number of clusters was calculated (Bennett et al. 1991) and adjusted as the population of clusters from which the sample was drawn was small (Thrusfield 1995). An estimated prevalence (P) of NSF STEC O157:H7 was set at 50% (Thrusfield 1995), the intracluster correlation coefficient (ρ) at 0.2 (Otte and Gumm 1997), and the significance level at 5%. The average number of cattle in a cluster was found to be five during a preliminary survey and thus used in calculation of the number of clusters. One hundred cattle-keeping households, who kept 446 cattle in total, were randomly selected from a sampling frame of 367 households and matched to their respective non-cattle–keeping neighbors living within a radius of 100 meters. Face-to-face interviews with household leaders using a structured questionnaire were used to gather information regarding cattle and manure management. Further information was collected by observations made around the households. Stools from one member of a cattle-keeping household involved in cattle and manure management and any member from a non-cattle–keeping household were collected early in the morning following prior notification and provision of a stool collection container the preceding evening. Water and soil samples were collected from the household the same morning that stool was collected. Soil samples (2–5 cm of top soil) were collected from five different locations of the household premises and mixed to form a pooled soil sample of about 100 grams. Surface water, such as borehole or pond, or any other available water source for human and/or cattle consumption at the household level was collected in sterile 250-mL bottles for isolation of E. coli. Individual fecal samples of 100–150 grams were collected by hand from the rectum using a disposable plastic glove from all cattle kept in a household. All samples were immediately placed in an insulated box with cooling elements and transported to the laboratory where bacteriological analysis was initiated. Some samples were kept overnight at 4–5°C for analysis the following day, with such samples being stored for a maximum of 24 h. A total of 200 human stool, 200 soil, 200 water, and 446 fecal samples from cattle were collected for E. coli analysis between December, 2010, and February, 2012.
Isolation of E. coli
One gram of stool, soil, and feces and 1 mL of water sample were mixed with 4 mL of normal saline (0.9% NaCl) to form a sample suspension. One milliliter of the sample suspension was enriched by incubation at 37°C for 24 h in 4 mL of brain infusion broth (CM0225, Oxoid, Hampshire, UK). Sample enrichments were subcultured onto Sorbitol MacConkey agar (CM0813, Oxoid) supplemented with cefexime-tellurite (SR0172E, Oxoid) and incubated at 37°C for 24 h. Medium sized, round, smooth, colorless (NSF) colonies were checked for purity after subculture onto blood agar plates and then tested for lactose fermentation on MacConkey agar (M082-500G, Himedia, Mumbai, India), β-glucuronidase and β-galactosidase activities on Brilliance E. coli agar (CM1046, Oxoid), and tryptophanase activity in the indole test.
All NSF E. coli isolates were subsequently tested for the presence of O157 somatic antigen by agglutination test using a Wellcolex E. coli O157 kit (Remel Europe Ltd., UK) according to manufacturer's instruction.
Detection of diarrheagenic E. coli
Multiplex PCR for diarrheagenic E. coli was done for all NSF E. coli isolates using a diarrheagenic E. coli (DEC) PCR kit (SSI Diagnostica, Hillerød, Denmark, 2011) to check for presence of the following genes: intimin gene (eae) with amplicon size 377 bp; verocytotoxin 1 (vtx1) with amplicon size 260 bp; verocytotoxin 2 (vtx2) with amplicon size 420 bp; heat-stable enterotoxin, human variant (estA-human) with amplicon size 151 bp; heat-stable enterotoxin, porcine variant (estA-porcine) with amplicon size 160 bp; heat-labile enterotoxin (eltA) with amplicon size 479 bp; invasive plasmid antigen (ipaH) with amplicon size 647 bp; and 16S rDNA genes with amplicon size 1062 bp. The latter gene served as a positive PCR internal control. Template DNA preparation, PCR master mix and PCR reaction volume were prepared according to the manufacturer's instructions. PCR amplification was run in a thermocycler at the following conditions: Initial denaturation at 95°C for 2 min followed by 35 cycles of denaturation at 94°C for 50 s, annealing at 62°C for 40 s, and extension at 72°C for 50 s. The final extension was done at 72°C for 3 min. For each sample and the controls, 18 μL of PCR product was run in separate wells on 2% agarose gel at 125 V for 1 h followed by staining in ethidium bromide (SSI-Diagnostica 2011). STEC isolates were subtyped by PCR for vtx1 and vtx2 genes (Scheutz et al. 2012). Vero cell cytotoxicity assay was performed for all isolates positive for virulence genes to test for cytopathic effects on Vero cell monolayers (Scheutz 1997).
Strains found to harbor virulence genes in the DEC PCR were confirmed by dot-blot DNA hybridization and tested for additional virulence genes at the Statens Serum Institut, Denmark. DNA probes included vtx1, vtx2, eae, ehxA, EAF, bfpA, saa, astA, and vtx2f. Isolates were allowed to grow on the nylon membrane positioned on top of an agar plate. The colonies were lysed, denatured, and neutralized using standard conditions and then hybridized (Scheutz 1997).
Serotyping of diarrheagenic E. coli
O and H antigen serotyping was performed according to standard procedures at the Statens Serum Institut (Ørskov and Ørskov 1984; SSI 2011). In summary, boiled cultures of isolates were tested against pooled O antisera and single O antisera of respective pools. Infusion broth inoculated with a colony of isolate was incubated at 37°C for 24 h. Broth cultures were boiled at 100°C for 1 h and left to cool on the table for 1 h before use. Eighty microliters of pooled O antisera was mixed with an equal amount of boiled culture in microtiter plates and incubated in humid atmosphere at 50°C overnight. A boiled culture positive for pooled O antisera was tested with an equal amount of single O antisera and incubated at 50°C overnight. A positive reaction—agglutination covering the bottom of microtiter well—was read against artificial light on a black background.
For H antigen typing, one colony from agar medium was inoculated in semisolid medium and incubated at 37°C. Motile cultures were passed to another semisolid medium using a Pasteur pipette and incubated overnight at 37°C. Cultures were further passed to nutrient beef broth, incubated at 37°C for 6 h in a roller device, and fixed with formaldehyde 0.5%. Pooled H antiserum (180 mL) was mixed with an equal amount of fixed culture in a glass tube. The reaction was read after 2 h incubation of the mixture at 50°C. Positive fixed cultures were tested with an equal amount of single H antisera of the respective H pool. A positive agglutination reaction was fluffy.
Antimicrobial susceptibility test
Antimicrobial susceptibility testing of all NSF E. coli isolates was determined by the disc diffusion method according to standards of the Clinical Laboratory Standard Institute (CLSI 2008). The following discs (Oxoid, UK) were used: Amoxillin (30 μg), ampicillin (10 μg), sulfamethazole/trimethoprim (25 μg), tetracycline (30 μg), ciprofloxacin (5 μg), gentamicin (10 μg), streptomycin (10 μg), and cefotaxime (30 μg). Fresh colony suspension, equivalent to 0.5 McFarland standards, was tested against the antimicrobial discs on Mueller–Hinton agar, and the diameters of the zone of complete inhibition were read after 18 h of 37°C incubation. Isolates were also tested for presence of extended spectrum β-lactamase (ESBL) genes for the TEM, SHV, OXA-10 group, and the CTX-M group 1, 2, 8, and 9 enzymes by PCR, as previously described (Hansen et al. 2012). The O25-ST131 clone allele-specific PCR for the paB gene was performed as previously described (Clermont et al. 2009).
Ethical approval
The study was approved by the Tanzania National Institute for Medical Research (NIMR) Ethical Board (NIMR/HQ/R 8a/Vol. IX/927). Confidentiality of the study outcome and all conditions for research approval were observed throughout the study
Results and Discussion
NSF E. coli
From the total of 1046 samples analyzed, 143 (13.7%) yielded NSF E. coli, including 95 (9.0%) isolates from cattle feces, 33 (3.2%) from human stool, eight (0.8%) from soil, and seven (0.7%) from water samples. Only one isolate was selected from each sample. Four isolates (4.2%) from cattle were positive for O157 somatic antigen. Traditionally, the inability of E. coli O157:H7 to ferment sorbitol has been used to differentiate them from other strains. The use of sorbitol MacConkey agar to screen for STEC O157:H7 is reported to miss out some nonmotile VTEC O157 strains because they can ferment sorbitol (Karch and Bielaszewska 2001); however, recovery of STEC (including non-O157), EPEC, and A/EEC in the present study (Table 1) illustrates that there is a broad range of diarrheagenic E. coli that do not ferment sorbitol.
VTEC, Vero cytotoxin–producing E. coli; A/EEC, attaching and effacing E. coli; EPEC, enteropathogenic E. coli.
Multiplex DEC PCR of all NSF E. coli revealed that 11 isolates (7.7%) carried at least one virulence gene (Fig. 1). The proportion of cattle carrying NSF diarrheagenic E. coli was 10.5%, whereas that of water samples positive for NSF diarrheagenic E. coli was 14.3%. An amplicon was produced for 16S rDNA in all tested isolates except for the negative control, confirming that the PCR was done correctly. The results from the multiplex DEC PCR, DNA hybridization, and O:H serotyping of the NSF E. coli are shown in Table 1. Isolates contained either one or a combination of vtx1, vtx2, eae, ehxA, EAF, bfpA and astA genes. Four isolates carried both eae and vtx2 genes, two isolates carried vtx1 and vtx2, and two isolates carried EAF and bfpA genes. Six out of 11 diarrheagenic E. coli isolates had cytotoxic effects on Vero cells (Table 2). The O157:H7 was the most commonly isolated serotype (four isolates), while other isolates were O113:H21, O142:H34, O+:H−, and O+:H16. One STEC isolate was nonmotile and two isolates had an O antigen that could not be determined (Table 1). The vtx subtypes were dominated by vtx2c (five out of seven isolates) (Table 2). Three bovine isolates had more than one subtype of the vtx gene, while one isolate contained three vtx subtype genes (Table 2).
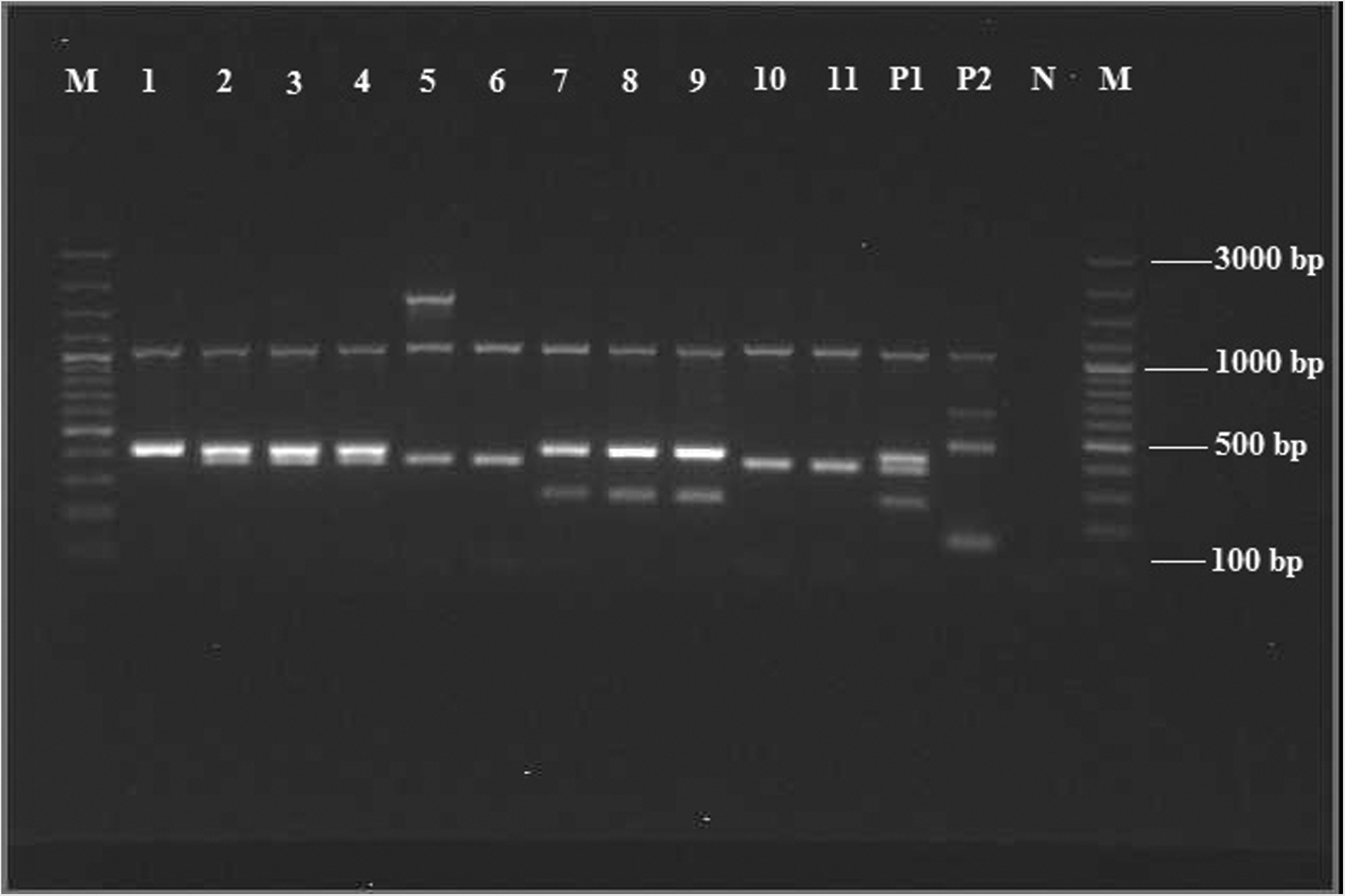
Multiplex DEC PCR for non-sorbitol–fermenting E. coli isolates. Lanes M, molecular weight size marker (100-bp plus DNA ladder); lane 1, BMZU001 (vtx2); lane 2, BMKB070 (vtx2 and eae); lane 3, BMKB068 (vtx2 and eae); lane 4, BMKB069 (vtx2 and eae); lane 5, BKIN069 (eae); lane 6, BKIH015 (eae); lane7, BKIH040(2) (vtx1 and vtx2); lane 8, BKIH101 (vtx1 and vtx2); lane 9, BBIG020(1) (vtx1 and vtx2); lane 10, BKIL032 (eae); lane 11, WKIL019(eae); lane P1, positive control for vtx2, eae and vtx; lane P2, positive control for ipaH, eltA and estA; lane N, negative control.
Occurrence of genes vtx1, vtx2, eae, ehxA, and astA in different combinations and the cytopathic effect of isolate extracts on Vero cells suggest that these isolates can cause disease in humans (Boerlin et al. 1999, Blanco et al. 2003, Caprioli et al. 2005). Different vtx2 subtypes were found in the current study, a finding also reported earlier (Scheutz et al. 2001). Virulence genes eae and vtx2 have been more often associated with serotypes reported from human diseases than vtx1 (Boerlin et al. 1999). Moreover, specific subtypes of vtx2 have been associated with HUS cases, whereas other subtypes seem to be most commonly associated with uncomplicated diarrhea (Friedrich et al. 2002). Our recovery of vtx1a, vtx2b, vtx2c, and vtx2d in different combinations and in some isolates in combination with the eae gene shows that inadequate management of cattle manure and associated human exposures may represent health risks for people living in urban areas with livestock keeping.
Prevalence of STEC
The prevalence of STEC O157:H7 in cattle was 0.9% [95% confidence interval (CI) 0.29–2.15], whereas the prevalence of STEC in cattle was 1.6% (95% CI 0.69–3.08). The overall prevalence of diarrheagenic E. coli in cattle feces was 2.2% (95% CI 0.99–3.67) and 0.5% (95% CI 0.025–2.44) in water. Each individual diarrheagenic E. coli isolate came from a different source, in which case nine isolates originated from different cattle and herds and one isolate was isolated in a water sample in a household where diarrheagenic isolates could not be recovered from either human stool, cattle feces, or soil samples. There may be little transmission within clusters, but these scattered multiple sources increase the chance of pathogen transmission to humans, animals, and the environment in close vicinity to the clusters positive for diarrheagenic E. coli.
The low prevalence of NSF STEC O157:H7 found in cattle in Morogoro is similar to what has been reported in Kansas (1.3%) (Sargeant et al. 2000) and in Washington State for dairy cattle (0.28%), beef cattle on pasture (0.71%), and beef cattle in feedlots (0.33%) (Hancock et al. 1994). However, higher occurrence has been reported elsewhere in South Africa, where 5.4–20% of cattle were positive for STEC O157:H7 (Ateba and Bezuidenhout 2008). The prevalence of 1.6% STEC in cattle in the present study is comparable to what has been reported in Spain in adult healthy cattle (2.5%), but is lower than that in calves (7.9%) and heifers (20.2%) in the same country (Orden et al. 2002). The sensitivity of diarrheagenic E. coli isolation may have been negatively affected by the amount of sample and the type enrichment media used. In the present study, 1 gram of sample was inoculated in 4 mL of nonselective enrichment medium. However, the method used is comparable to a study by Sargeant et al. (2000), who enriched 10 grams of sample in 90 mL of enterohemorrhagic E. coli (EHEC) enrichment broth and reported a prevalence of NSF STEC of 1.3%. On the contrary, 20 grams of the sample enriched in 180 mL of modified E. coli broth yielded a prevalence of 11.1%% NSF STEC O157:H7 (Heuvelink et al. 1998).
There are no previous records for Tanzania and neighboring countries of the prevalence of STEC O157:H7 in cattle. Thus, the estimated prevalence of STEC O157:H7 was set at 50% to maximize the sample size when the sample size was calculated (Thrusfield 1995). However, non-O157 STEC has been isolated from both cattle and children in neighboring Uganda (Majalija et al. 2008), and a study on E. coli O157:H7 in human patients with diarrhea in Morogoro, Tanzania, found that 0.7% of strains harbored vtx1 or vtx2 as well as the eae gene (Raji et al. 2008). It remains to be investigated whether EPEC strains in cattle feces and water sources as documented in this study may be associated with human diseases, e.g., gastroenteritis in children due to EPEC infection that has been reported in Dar es salaam, Tanzania, where livestock is also kept in urban areas (Moyo et al. 2011). Our results document for the first time that STEC (including E. coli O157:H7), EPEC, and A/EEC strains have been found in cattle and water sources in Tanzania. However, if the use of sorbitol MacConkey agar to recover NSF E. coli can detect diarrheagenic E. coli at this rate, questions arise about the magnitude of the prevalence of sorbitol-fermenting diarrheagenic E. coli and sorbitol-fermenting O25:H4, ST131 in this environment.
Antimicrobial resistance
The diarrheagenic E. coli isolates were all susceptible to amoxillin, ampicillin, sulfamethazole/trimethoprim, tetracycline, ciprofloxacin, gentamicin, streptomycin, and cefotaxime. However, a nondiarrheagenic isolate from human (O25:H4) (Table 1) tested positive as O25b-ST131 by the clone allele-specific PCR for the paB gene, which is identical to the worldwide ESBL-producing clone and was positive for the CTX-M-gr. 1 gene. ESBL variant TEM-63 and CTX-M-15 have previously been isolated from blood cultures of patients in Dar es Salaam, Tanzania (Blomberg et al. 2005), and the isolation of O25:H4, ST131 from the stool of a healthy individual in the current study gives another dimension of concern of resistant gene spread among the population.
Cattle and manure management practices
Cattle and manure management practices for diarrheagenic E. coli–positive herds are summarized in Table 3. In general, cattle at the study sites and elsewhere in periurban areas of Tanzania roam around, and different herds are allowed to mix while foraging during daytime and are then housed at night within residential areas. The manure accumulated overnight is collected by bare hands, spades, and wheelbarrows and disposed of as fresh manure in and around residential areas in both households with diarrheagenic E. coli–positive and −negative herds. The effluent from the animal house was indiscriminately discharged and leached into the surrounding land in all nine (9%) households with cattle positive for diarrheagenic E. coli and in 71 (71%) households with negative herds. In all nine (9%) households with cattle positive for diarrheagenic E. coli, humans and cattle were found to share common water sources for drinking purposes compared to 51 (51%) households with diarrheagenic E. coli–negative herds (Table 3). However, the ratio of diarrheagenic E. coli–positive to −negative herds in each category did not show evidence of a relation between cattle and manure management practices and isolation of diarrheagenic E. coli
During grazing, cattle may have contact with other animals, including wildlife. The indiscriminate defecation on grazing land and at household premises increases exposure and spread of pathogens between animals, humans, and the environment, e.g., children playing around the household perimeter are likely to have high exposure to animal manure through contact with contaminated soil. Manure effluents are also discharged from cattle houses, resulting in further contamination of the surrounding land, with cattle keepers and their household members being at increased risk of exposure to diarrheagenic E. coli. At the study sites, cattle and households often share sources of surface water, such as rivers and ponds, especially during the dry season when these are important common sources of drinking water. Close contact between human and livestock as documented in this study has been reported to result into sharing of nonpathogenic commensal bacteria between humans and different animal species (Rwego et al. 2008). Given such interactions, the presence of pathogenic microbes can lead to either human or animal infection or environmental contamination. For instance, outbreak of enteric infectious diseases, including those due to E. coli O157:H7, have been reported among people on farm visits in the United States who had direct contact with manure from infected cattle (Crump et al. 2002, Smith et al. 2004). However, it is uncertain to what extent the occurrence of diarrheagenic E. coli found in cattle and in water in the present study represents real human health hazards. Therefore, there is a need for further research to quantify possible risk factors for pathogen transmission from animals or environment to animal keepers and neighboring community in the study area and recommend appropriate measures to safeguard public health.
Footnotes
Acknowledgments
The authors wish to thank the Ministry of Foreign affairs of Denmark through the Danish International Development Agency (Danida) for financial support to the project “Opportunities and challenges in peri-urban livestock farming in Tanzania” (P6-08-Tan). The excellent collaboration between Sokoine University Agriculture (Tanzania) and University of Copenhagen (Denmark) is highly appreciated. We acknowledge participation and project support of cattle keepers and extension staff in urban and peri-urban areas of Morogoro. The dedicated technical assistance in this study by Pernille Gymoese, Frank Hansen, and Susanne Jespersen is greatly appreciated.
Author Disclosure Statement
No competing financial interests exist.