
Abstract
Toxocara canis is a zoonotic nematode parasite that can be transmitted to humans by food or water contaminated with T. canis eggs from infected dog feces. High-pressure processing (HPP) is a useful alternative to thermal treatments to eliminate pathogens from foods. Most of the research on HPP has focused on prokaryotes, but little is known about its effects on eukaryotic organisms. We evaluated the ability of HPP to affect embryonation of T. canis eggs to test the hypothesis that HPP treatment can delay development of T. canis eggs. Efficacy of HPP was determined by using an embryonation assay on T. canis eggs from naturally infected puppies. For each treatment, 2500 T. canis eggs in tap water were placed in sealable plastic bags and subjected to 138–400 megapascals (MPa; 1 MPa=10 atm=147 psi) for 60 s in a commercial HPP unit. We found that treatment with 300 or 400 MPa for 60 s killed 100% of eggs using embryonation as the standard. Treatment with 250, 241, and 207 MPa was less effective and killed 80%, 56%, and 8% of eggs, respectively. Results from this study suggest that HPP may be a useful treatment to protect foods from T. canis contamination.
Introduction
Toxocara canis,
High-pressure processing (HPP) is a nonthermal alternative for removing nonspore-forming bacteria from various food products (Flick 2003). Foods treated with HPP exhibit extended shelf life and sensory features (flavor, color, etc.) or are minimally affected or unaffected by HPP.
Research on HPP has focused on prokaryotic pathogens, but few studies have investigated the effects of HPP on metazoans. A study on Trichinella spiralis revealed that 8-week-old larvae are killed by pressures greater than 200 megapascals (MPa; 1 MPa=10 atm=147 psi) (Ohnishi et al. 1992, 1994). Gamble et al. (1998) showed that pressures of 55–60 MPa did not kill Tr. spiralis larvae in pork tenderloin or diaphragm. Porto-Fett et al. (2010) determined that Tr. spiralis larvae in pig masseter and pork genoa salami were inactivated when subjected to 483 MPa or 600 MPa by HPP. Larvae of Anisakis simplex were killed using lack of motility as an indicator of larval death after treatment with 200 MPa for 10 min at 0–15°C (Molina-Garcia and Sanz 2002). Dong et al. (2003) demonstrated that 100% An. simplex larvae in arrowtooth flounder and king salmon were killed at 414 MPa for 30–60 se, 276 MPa for 90–180 s, and 207 MPa for 180 s. Merwad et al. (2011) showed that ova of Hymenolepis diminuta, the rat tapeworm, were killed at pressures of ≥300 MPa for 60 s using an egg hatching assay to determine viability. Eggs of the canine whipworm Trichuris vulpis were inactivated and failed to embryonate after subjection to≥400 MPa for 60 s (Rosypal et al. 2011). High-pressure treatment of Ascaris suum eggs, the pig roundworm, for ≥60 s at 241 MPa or 10–30 s at 276 MPa rendered eggs nonviable (Rosypal et al. 2007). Rosypal et al. (2011) demonstrated that HPP treatment caused a delay in embryonation of Tri. vulpis and A. suum eggs 4–16 days compared to control eggs (Rosypal et al. 2011).
Produce contaminated with T. canis eggs is a source for transmission of the parasite to humans (Slifko et al. 2000). Toxocara eggs have been found on radishes and carrots (Vazquez Tsuji et al. 1997) as well as leafy vegetables, such as lettuce and parsley (Avcioglu et al. 2011). The potential for HPP to control pests on produce as a quarantine method has recently been investigated (Neven et al. 2007, Velazquez et al. 2010). These studies examined the effects of HPP on eggs and larvae of insect pests, but it is possible that HPP could also be applicable as a means of preventing food-borne Toxocara infections. Currently, there is nothing known about the susceptibility of T. canis to HPP. The purpose of the present study was to determine the effects of high hydrostatic pressure on embryonation of T. canis eggs.
Materials and Methods
T. canis eggs
T. canis eggs were collected from the feces of naturally infected puppies housed at the Montgomery County Human Society, Christiansburg, VA. Fecal samples were collected from these puppies and kept refrigerated at 4°C until used. Fecal material was mixed with Sheather sugar solution, filtered through a tea strainer into 50-mL centrifuge tubes, and centrifuged for 10 min to concentrate eggs by flotation. T. canis eggs in the top 2–3 mL from the surface were collected. Isolated eggs were washed in tap water and counted, and their volume was adjusted to a concentration of 5000 eggs/mL. T. canis eggs were stored at 4°C in tap water until used for the present study.
HPP of T. canis eggs
For each HPP treatment, 2500 isolated eggs in 0.5 mL water were placed in sealable pouches (Kapak Sealpak pouches, Minneapolis, MN). The pouches were compressed to force out any air, and they were sealed with a sealing machine (Shop Sealer FS-315, Fugi Impulse Vietman Co. Ltd., Sai Gon-Linh Trung EPZ, Japan). To prevent contamination, the bags containing T. canis eggs were placed in a second leak-proof, resealable bag and vacuum-sealed. The double-bagged, vacuum-sealed bags containing 2500 eggs each were subjected to high pressures equal to 0, 138, 207, 241, 250, 300, and 400 MPa for 60 s each using a commercial HPP unit (Quintus Food Press QFP 35L-600 Model, Flow International Corporation [Avure Technologies], Kent, WA) with a 7XS-6000-intensifier pump and maximum operating pressure of 600 MPa. The HPP unit was installed and operated at Virginia Tech's Department of Food Science and Technology.
Preparation of cultures and examination of treated eggs
After treatment, each sample was mixed in 4.5 mL of 0.01 N sulfuric acid and placed individually in to a 25-cm2 vented-cap tissue culture flask. The flasks were placed on a rotating mixer platform (Thermolyne Platform Vari Mix, VWR Scientific Products, Radnor, PA) and rotated continuously at room temperature for 35 days. The 25-cm2 culture flasks were examined using an inverted microscope (Zeiss Invertoskop), and notes on development of 25 (days 1, 2, 3, 4) or 50 (days 7, 8, 10, 17,18, 21, 22, 23, 27, 28, 32, and 35) eggs in each flask were recorded. Embryonation percentages were calculated from these numbers. The first day an egg containing a motile larvae was observed was designated as the day of embryonation.
Results and Discussion
The results of HPP treatment on T. canis eggs are presented in Table 1. No obvious structural damage was observed in any of the eggs from any of the treatment groups (Fig. 1). Treatment with 300 or 400 MPa for 60 s was 100% effective in killing eggs using embryonation as the standard. These eggs appeared to be structurally normal and looked like freshly excreted eggs (Fig. 1B). Treatment with 250 or 241 MPa was less effective, killing 80% and 56% of eggs, respectively. Treatment with 207 MPa had little effect and killed only 8% of eggs, whereas treatment with 138 MPa was no different from the control (Table 1). Treatment with 207, 241, and 250 MPa resulted in larvae being present 1, 3, and 10 days later, respectively, than the control. These findings suggest that T. canis eggs are slightly more resistant to HPP that are the eggs of A. suum.
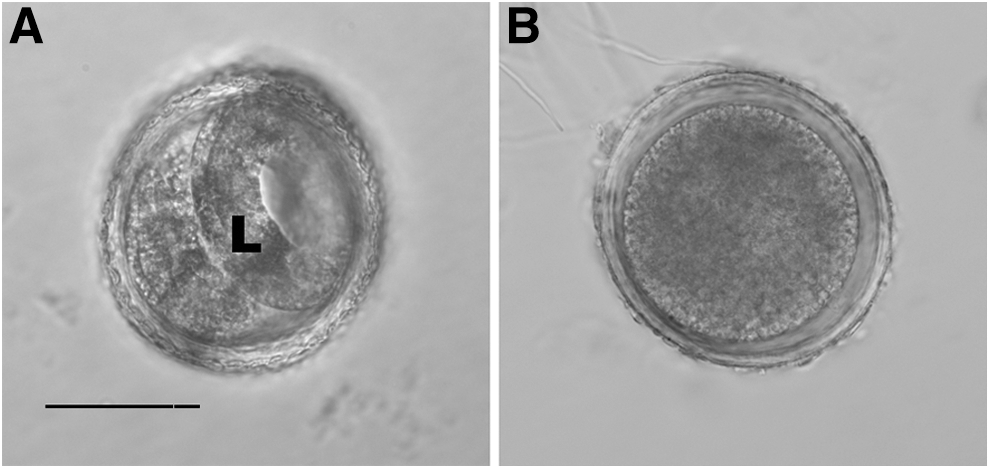
Eggs of T. canis examined 7 days after being exposed to high-pressure processing (HPP). Bar, 50 μm. (
MPa=megapascals (1 MPa=10 atm=147 psi).
Day motile larvae first observed.
Difference in number of days when larvae first observed in control versus culture treated with that MPa.
NA, Not applicable.
Rosypal et al. (2011) reported that embryonation of A. suum eggs was delayed by 4, 10, and 16 days for eggs treated with 207, 241, and 250 MPa, respectively, compared to nontreated control eggs. A. suum eggs treated with 207 MPa eventually embryonated to a similar percentage of embryonation values as controls and 138 MPa-treated A. suum eggs, but A. suum eggs treated with 241 or 250 MPa were always <5% embryonated. Rosypal et al. (2011) suggested that subpopulations of HPP-resistant eggs are present in A. suum. A similar phenomenon was observed in T. canis eggs in this study.
We were not able to measure the infectivity of T. canis larvae produced in HPP-treated eggs that survived treatment. Motility suggests that the larvae are viable but does not demonstrate that they are infective. The exact mechanism of inactivation in HPP-treated eggs is unclear; however, treatment with HPP affects three-dimensional protein shape and denaturation by impacting hydrogen bonds (Hayakawa et al. 1996). Radiation has been used to attenuate nematode parasites and to produce live vaccines for use in veterinary medicine (Boag et al. 2003). Results from this study suggest that HPP may also be used as an alternative to radiation as a means to attenuate nematode parasites for live vaccine development. It would be interesting to examine the ability of larvae produced by megapascal treatment of eggs to develop in dogs and paratenic hosts. It is possible that the larvae may be infective and produce an immune response but not develop to patency (in dogs) or persist in paratenic hosts.
Footnotes
Acknowledgments
The contribution of ACR was supported in part by the Faculty Summer Retooling Program funded by the Historically Black University Undergraduate Program at Johnson C. Smith University.
Author Disclosure Statement
No competing financial interests exist.