
Abstract
Birds serve as reservoirs for at least 10 arthropod-borne viruses, yet specific immune responses of birds to arboviral infections are relatively unknown. Here, adult House Sparrows were inoculated with an arboviral alphavirus, Buggy Creek virus (BCRV), or saline, and euthanized between 1 and 3 days postinoculation. Virological dynamics and gene expression dynamics were investigated. Birds did not develop viremia postinoculation, but cytopathic virus was found in the skeletal muscle and spleen of birds 1 and 3 days postinoculation (DPI). Viral RNA was detected in the blood of BCRV-infected birds 1 and 2 DPI, in oral swabs 1–3 DPI, and in brain, heart, skeletal muscle, and spleen 1–3 DPI. Multiple genes were significantly upregulated following BCRV infection, including pattern recognition receptors (TLR7, TLR15, RIG-1), type I interferon (IFN-α), and type II interferon (IFN-γ). This is the first study to report avian immunological gene expression profiles following an arboviral infection.
Introduction
A
Specific cellular mechanisms of avian immune responses to viral infections have been well documented in domestic poultry and ducks, with the majority of studies being focused on avian immune responses to Newcastle disease virus, Marek's disease, and influenza virus (Schat and Xing 2000, Seal et al. 2000, Barber et al. 2010). However, no studies exist that have explored the critical host responses involved in avian alphavirus infections.
Pattern recognition receptors (PRRs) are conserved receptors that recognize pathogen-associated molecular patterns (PAMPs). Several PRRs have been implicated in the avian response to viral infections, including Toll-like receptors (TLRs) TLR2, TLR3, TLR4, TLR7, TLR9, and TLR15 (Kawai and Akira 2006, Kaiser 2010, Nang et al., 2011, Boyd et al. 2012), and retinoid-inducible gene 1 (RIG-1), a cytoplasmic PRR (Loo et al. 2008). In mammalian alphavirus infections, TLR3, TLR7, and TLR9 stimulate antiviral inflammatory pathways and interferon production (Esen et al. 2012, Neighbours et al. 2012). Type I interferons (IFN-α and IFN-β) have been shown to play an essential role in the control of alphavirus infections such as VEEV (Grieder and Voge 1999), SINV (Ryman et al. 2000, Ryman et al. 2007, Zhang et al. 2007, Ryman and Klimstra, 2008), and WEEV (Wu et al. 2007). In mammals, severe alphavirus-induced disease is usually characterized by over production of proinflammatory cytokines, often called systemic inflammatory response syndrome (SIRS), which may be followed by neuroinvasion and death (Klimstra et al. 1998, Ryman and Klimstra 2008).
The objectives of this study were to determine specific virological parameters (viremia and muscosal shedding intensity and duration, and viral dissemination patterns in tissues) and expression of selected immune function genes in House Sparrows following infection with two different lineages of BCRV (lineages A and B). On the basis of previous field work conducted on BCRV infections in House Sparrows and studies conducted on mammalian alphavirus infections, we predicted that adult House Sparrows would develop mild BCRV infections, and predicted upregulation of key antiviral genes such as IFN-α and virus-specific PRRs postinfection.
Materials and Methods
Experimental inoculation
Eighteen adult House Sparrows were collected near Council Bluffs, Iowa, using mist nets. Birds were then transported to a captive animal facility where they were placed in wire cages in a temperature-controlled room and 12-h illumination. Starting on day 5 postcapture, background blood samples (50 μL) were collected via jugular or brachial venipuncture. The samples were diluted with BA-1 diluent and centrifuged at 6000× g for 5 min; the serum was stored at −80°C until it was tested for BCRV-specific antibodies according to Fassbinder-Orth et al. (2013). All 18 birds were negative for BCRV-specific antibodies. Background oral pharyngeal swabs were collected in complete Eagle minimal essential medium (EMEM) containing heat-inactivated 10% fetal bovine serum (FBS), penicillin (100 U/mL), streptomycin (100 μg/mL), and amphotericin B (0.25 μg/mL), and stored at −80°C until further use. House Sparrows were randomly divided into three treatment groups (BCRV-A, BCRV-B, or control, n=6/group). Birds were inoculated subcutaneously in the pectoral region 8 days postcapture with a 100-μL solution containing 3.5 log10 plaque-forming units (PFU) of BCRV lineage A (BCRV-A), BCRV lineage B (BCRV-B), or a buffered saline solution serving as a negative control. BCRV used in the inoculation was obtained from naturally infected House Sparrow nestling serum samples, previously screened for lineage assignment (Padhi et al. 2008), and isolated through Vero cell culturing according to Fassbinder-Orth et al. (2013).
The course of the infection was monitored for 3 days postinoculation (DPI). Blood samples (50 μL) and oral pharyngeal swabs were collected daily from all birds postinoculation for use in the viral plaque assay and RT-PCR.
Tissue collection
Two House Sparrows per treatment were euthanized on days 1, 2, and 3 postinoculation by CO2 overdose. Immediately after euthanasia, necropsies were performed, and brain, heart, skeletal muscle, and spleen samples were collected for use in either viral plaque assays or gene expression assays. Tissues collected for plaque assays were placed in vials containing BA-1 diluent and immediately frozen at −80°C. Tissues collected for gene expression were immediately frozen in liquid nitrogen, then frozen at −80°C until further use. All procedures involving animals were approved by the Creighton University Institutional Animal Care and Use Committee under protocol 0904.
Plaque assay
Levels of cytopathic BCRV were assessed in blood, swab, and tissue samples using a Vero plaque assay, according to Fassbinder-Orth et al. (2013). Briefly, Vero cells in complete EMEM media were plated onto each well of a six-well plate and incubated for 3–4 days at 37°C and 5% CO2. Diluted samples were added to each well then incubated for 45 min. After incubation, 3 mL of an agar overlay was added for plaque visualization. Plates were incubated at 37°C and 5% CO2 and checked for cytopathic effect at 72 h and 96 h.
Selection of genes
Gene names, accession numbers, and gene functions are shown in Table 1. Gene sequences were produced for 14 genes, ranging from 361 to 675 bp in length. Sequences for three genes—β-actin, glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and BCRV E2 protein—were obtained from GenBank. Five housekeeping genes were chosen to improve the accuracy of target gene expression normalization (Radonić et al. 2004). Eight genes known to be important in the antiviral immune response were chosen, including six pattern recognition receptors (PRRs, TLR 2,3,4,7,15, and RIG-1) and two interferons (IFN-α and IFN-γ; Kawai and Akira 2006, Nang et al. 2011). The pleiotropic cytokine interleukin-10 (IL-10) was chosen for its role in suppressing vital T helper 1 (Th1) responses during viral infections, where it protects the host from inflammatory damage (Moore et al. 2001), but excessive levels can also facilitate viral persistence (Brooks et al. 2006). Matrix metallopeptidase-9 (MMP9) was chosen due to its role in inflammation during viral infections (Parks et al. 2004), and chemokine receptor CCR5 was included based on its involvement in viral entry and leukocyte trafficking (Glass et al. 2005). BCRV envelope protein gene was chosen to quantify viral loads in tissues.
Gene sequencing
RNA was extracted from adult House Sparrow tissues according to standard techniques using Qiagen's RNeasy kit (Qiagen Inc., Valencia, CA). RNA yields were determined via a NanoDrop Spectrophotometer (Thermo Scientific NanoDrop, Wilmington, DE). RT-PCR was performed using Qiagen's OneStep RT-PCR kit. Primers were designed based on available Zebra Finch (Taeniopygia guttata) mRNA sequences in GenBank. PCR products were visualized by gel electrophoresis using 1% agarose gels, purified using Qiagen's QIAquick PCR purification kit, and quantified on the NanoDrop Spectrophotometer. PCR products were sequenced (both forward and reverse sequencing primers were used to sequence each product) using Applied Biosystems 3130 Genetic Analyzer (Life Technologies, Grand Island, NY). Sequence analysis and alignment were performed using Applied Biosystems Sequencing Analysis software v.5.3.1, Applied Biosystems Sequence Scanner software v 1.0 (Life Technologies, Grand Island, NY), and MEGA 4.0.2 (Tamura et al. 2007).
mRNA quantification by multiplex branched DNA (bDNA) assay
Levels of mRNA were quantified from tissue lysates using the Quantigene Plex 2.0 Assay (Plex set 41215, Panomics/Affymetrix, Fremont, CA). Briefly, gene-specific bead-based oligonucleotide probes were developed for 12 target genes and five housekeeping genes by Panomics/Affymetrix. Diluted tissue lysates were analyzed using these fluorescent bead probes by the Luminex 100 xMAP system and BioPlex 5.0 software (Bio-Rad Laboratories, Hercules, CA). The geometric mean of three housekeeping genes (HPRT1, PGK1, and TFRC) was used to normalize levels of mRNA expression. β-Actin and GAPDH were not used as housekeeping genes in this study due to the relatively high and variable expression levels that were observed for these genes in this study.
RNA extraction and RT-PCR
To confirm the results of the bead-based gene expression assay for BCRV RNA presence in selected tissues, and to detect BCRV RNA in blood and swab samples, viral RNA was extracted from serum, swab, and tissue samples using the QIAamp Viral RNA Mini Kit (Qiagen Inc., Valencia, CA), and a 208-bp fragment of the E2 viral envelope protein-encoding region of the BCRV genome was amplified by RT-PCR according to Moore et al. (2007).
Statistical analyses
Samples from all BCRV-infected birds (A and B lineages) from all days postinfection (1–3 DPI) were analyzed together according to tissue type for the bead-based expression assay due to the lack of differences in virological parameters and gene expression recorded between the lineages and also among the days postinfection. Additionally, all control bird samples were pooled by tissue type and significant differences in gene expression were analyzed using a one-sample t-test (SYSTAT; Wilkinson 1999). A 1.5-fold change and p<0.05 were used as the cutoff values for significant gene expression changes, as suggested by Dalman et al. (2012).
Results
All 18 birds survived the duration of the experiment, and no birds showed signs of infection-related illness.
Viral detection
Neither viremia nor oral shedding of cytopathic virus was detected in any treatment group (Table 2). However, viral RNA was detected in the blood by RT-PCR in all BCRV-A– and BCRV-B–infected birds on 1 DPI, and all BCRV-A birds and 75% of BCRV-B birds 2 DPI. No viral RNA was detected 3 DPI for either BCRV treatment group. Additionally, viral RNA was detected in the oral swabs of 50% of all individuals (three birds per group) in BCRV-A and BCRV-B groups. Cytopathic BCRV was detected in the spleen and skeletal muscle of one bird for both BCRV-A and BCRV-B groups 1 DPI. No cytopathic BCRV was detected in brain or heart samples for any groups. BCRV RNA was detected by RT-PCR in all tissue samples for BCRV-A and BCRV-B infected birds except the heart of one individual 2 DPI for BCRV-A and 3 DPI for BCRV-B, and the brain of one individual 1 DPI for BCRV-A. BCRV RNA was detected by the multiplex bDNA assay in all tissues for all BCRV-infected birds 1–3 DPI (Table 2).
BBA, bead-based multiplex gene expression assay; NT, not tested.
Gene sequences and expression
PRRs
Virus-specific PRR expression was elevated in all tissue types sampled in BCRV-infected birds (Figs. 1 –4). Specifically, TLR7 expression was significantly higher in spleen (1.5-fold, p<0.0051), brain (5.9-fold, p<0.0001), and skeletal muscle (3.1-fold, p<0.01). TLR15 expression was significantly higher in brain (2.3-fold, p<0.003) and skeletal muscle (2.1-fold, p<0.0004). In the skeletal muscle, elevated levels of three additional TLRs were detected: TLR2 (4.0-fold, p<0.02), TLR3 (2.9-fold, p<0.0004), and TLR4 (5.7-fold, p<0.03). Last, RIG-1 expression was significantly higher in the spleen (1.5-fold, p<0.004), skeletal muscle (2.9-fold, p<0.03), and heart (1.5-fold, p<0.04) of BCRV-infected birds compared to control.
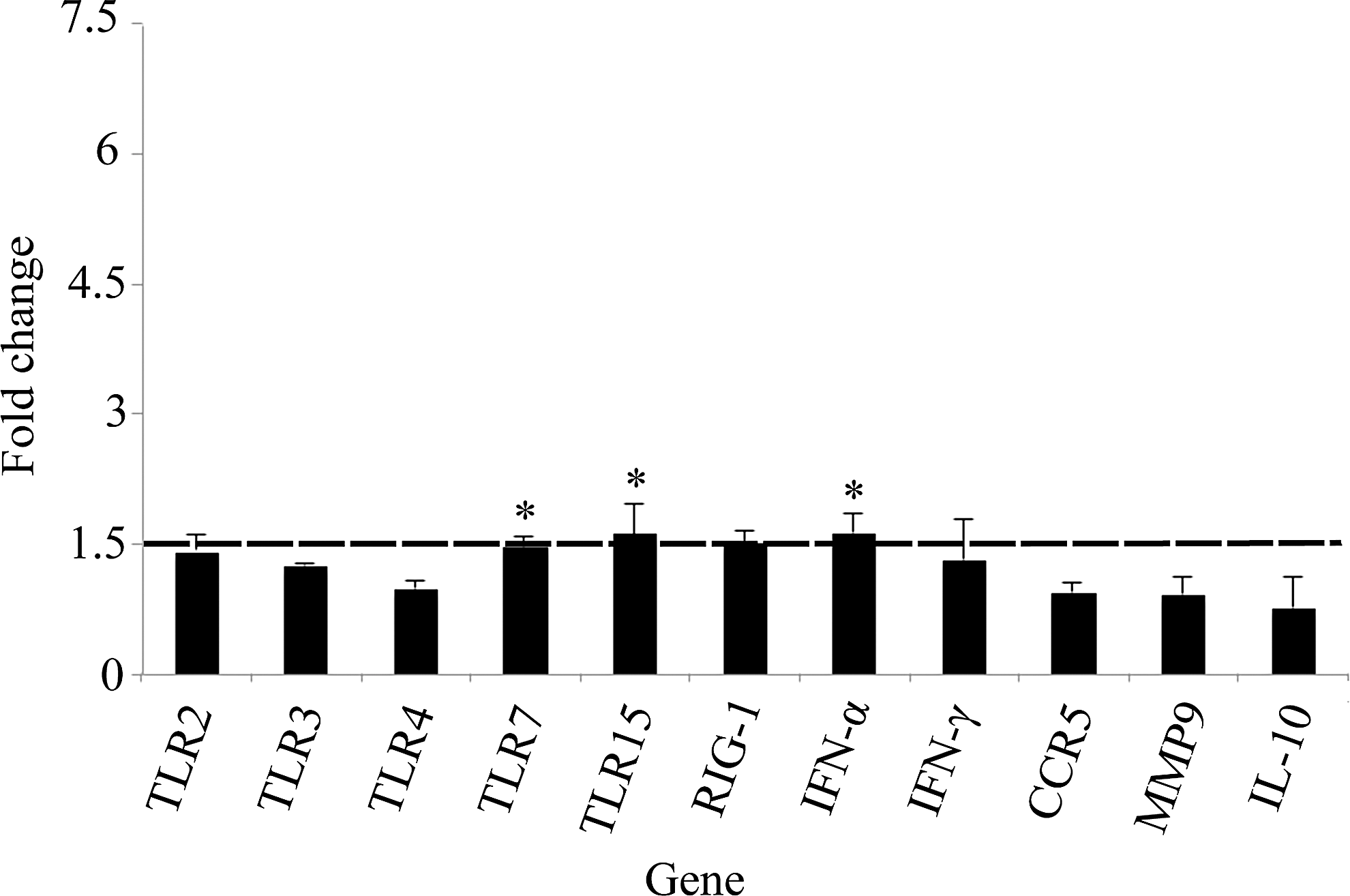
Fold change gene expression in spleen. Values shown are mean±standard error of the mean (SEM) fold change expression in Buggy Creek virus (BCRV)-treated individuals compared to controls. (*) Statistically significant increase in gene expression compared to control (fold change ≥1.5 and p<0.05). TLR, Toll-like receptor; RIG-1, retinoid-inducible gene 1; IFN-α, interferon-α; CCR5, C-C chemokine receptor type 5; MMP9, matrix metalloproteinase-9; IL-10, interleukin-10.
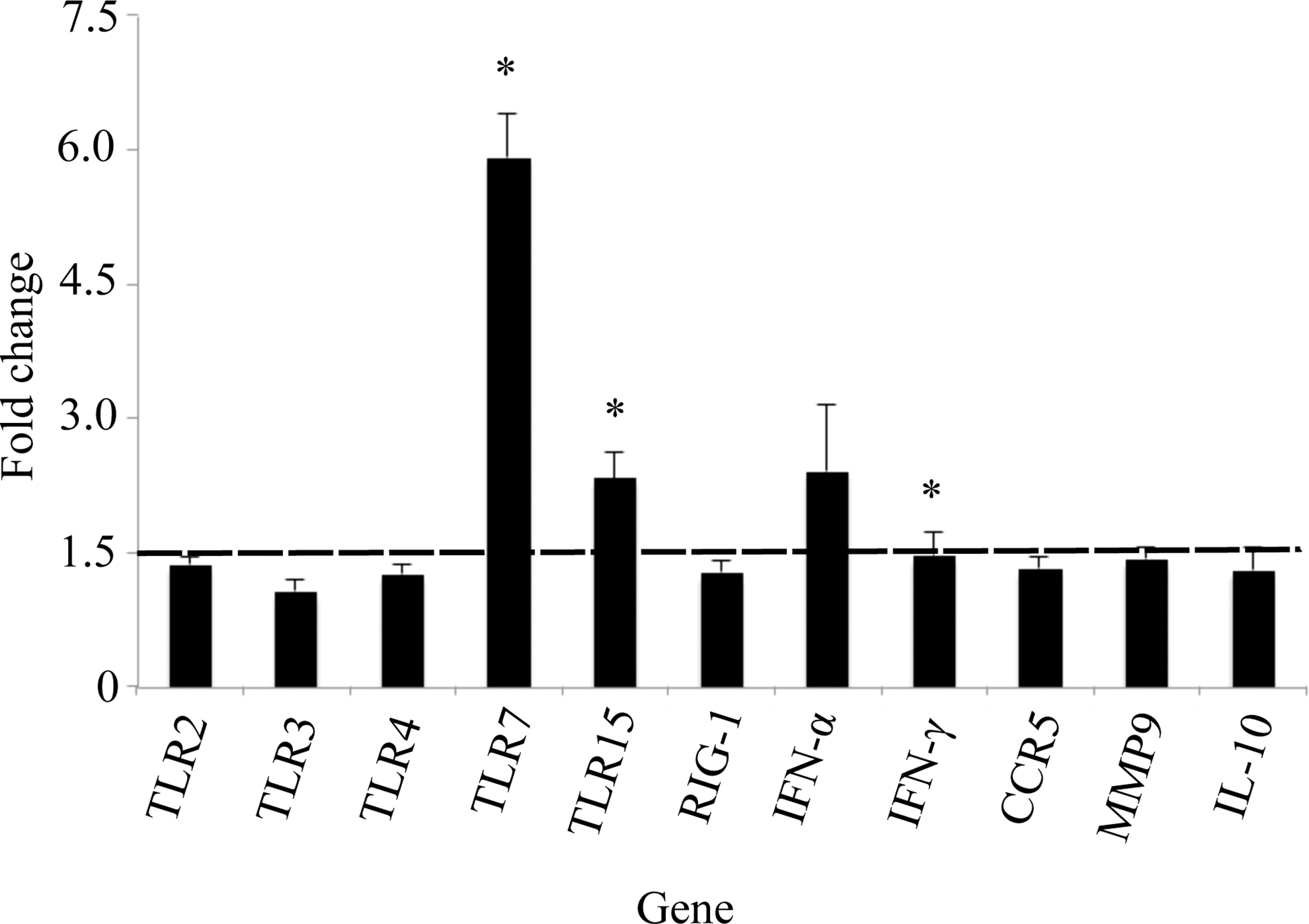
Fold change gene expression in the brain. Values shown are mean±standard error of the mean (SEM) fold change expression in Buggy Creek virus (BCRV)-treated individuals compared to controls. (*) Statistically significant increase in gene expression compared to control (fold change ≥1.5 and p<0.05). TLR, Toll-like receptor; RIG-1, retinoid-inducible gene 1; IFN-α, interferon-α; CCR5, C-C chemokine receptor type 5; MMP9, matrix metalloproteinase-9; IL-10, interleukin-10.
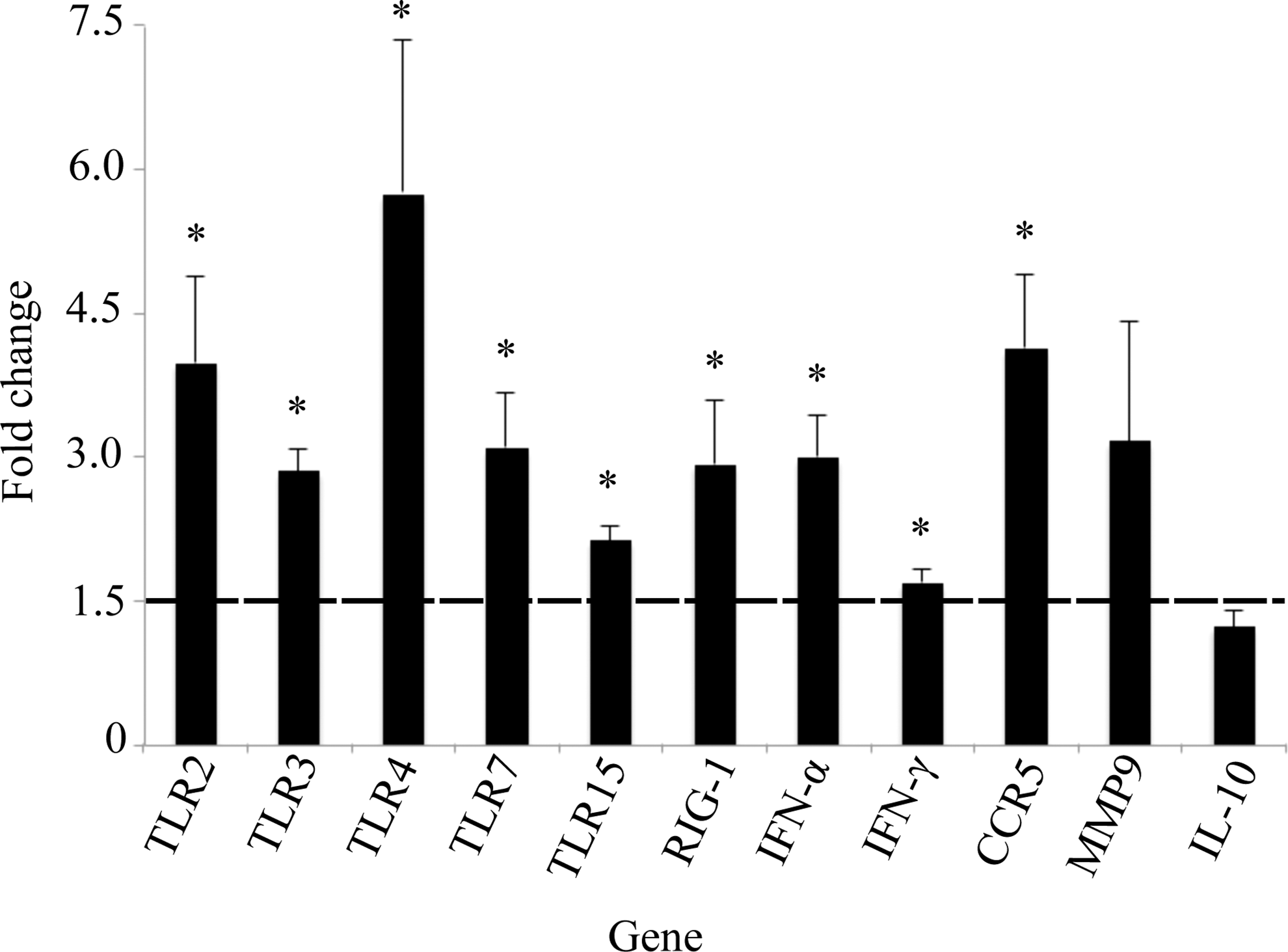
Fold change gene expression in the skeletal muscle. Values shown are mean±standard error of the mean (SEM) fold change expression in Buggy Creek virus (BCRV)-treated individuals compared to controls. (*) Statistically significant increase in gene expression compared to control (fold change ≥1.5 and p<0.05). TLR, Toll-like receptor; RIG-1, retinoid-inducible gene 1; IFN-α, interferon-α; CCR5, C-C chemokine receptor type 5; MMP9, matrix metalloproteinase-9; IL-10, interleukin-10.
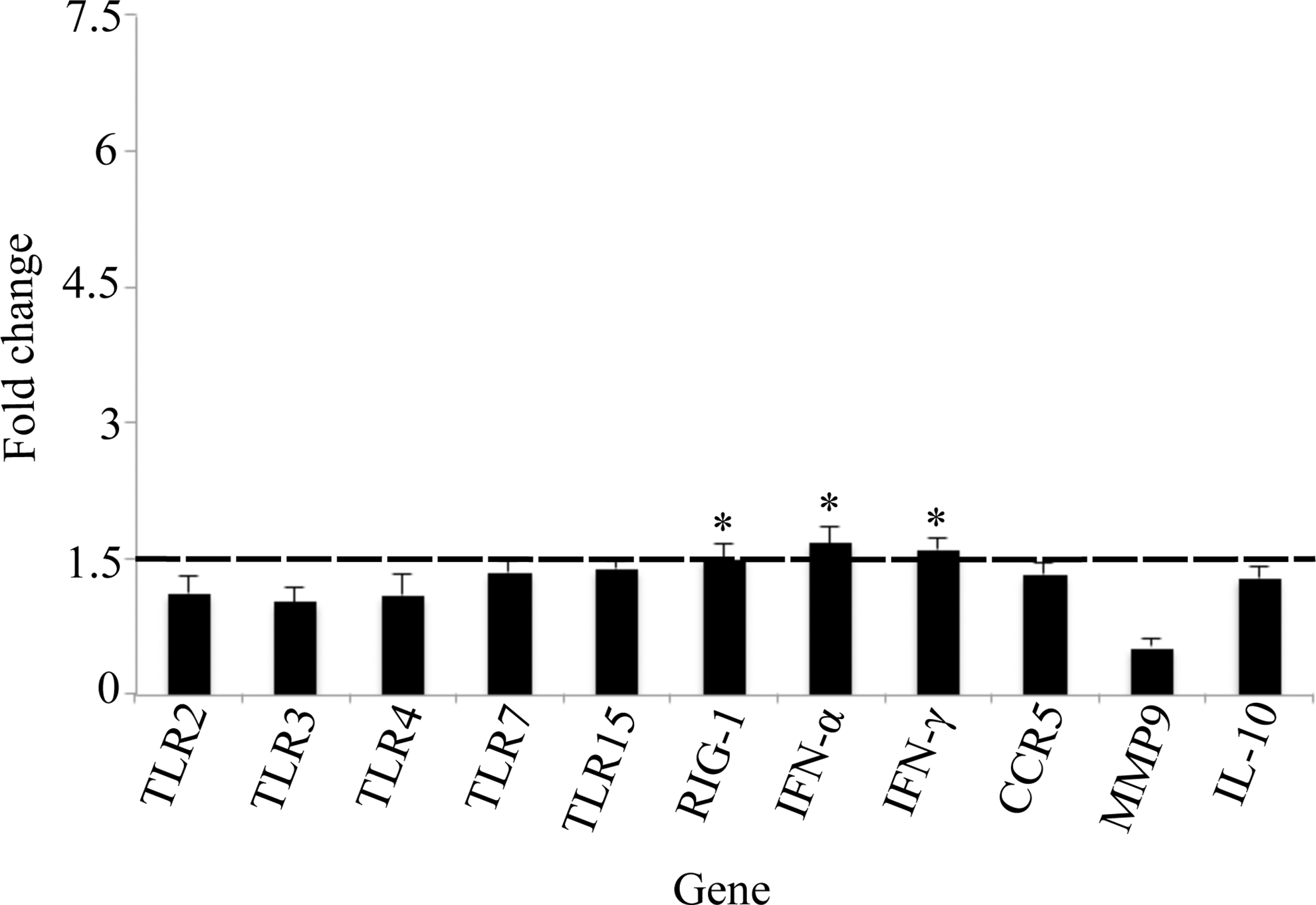
Fold change gene expression in the cardiac muscle. Values shown are mean±standard error of the mean (SEM) fold change expression in Buggy Creek virus (BCRV)-treated individuals compared to controls. (*) Statistically significant increase in gene expression compared to control (fold change ≥1.5 and p<0.05). TLR, Toll-like receptor; RIG-1, retinoid-inducible gene 1; IFN-α, interferon-α; CCR5, C-C chemokine receptor type 5; MMP9, matrix metalloproteinase-9; IL-10, interleukin-10.
Interferons
Levels of type I interferon (IFN-α) or type II interferon (IFN-γ) were significantly elevated in all tissue types sampled except the brain in BCRV-infected birds. Specifically, IFN-α expression was significantly higher in spleen (1.6-fold, p<0.03), skeletal muscle (3.0-fold, p<0.005), and heart (1.7-fold, p<0.01), whereas IFN-γ expression was significantly higher in the skeletal muscle (1.7-fold, p<0.004) and heart (1.6-fold, p<0.003).
Other genes
Expression of the chemokine receptor CCR5 was significantly increased in the skeletal muscle postinfection (4.1-fold, p<0.009), but was not elevated in other tissues. Additionally, IL-10 and MMP9 levels were not significantly elevated in any tissues postinfection.
Discussion
Virology
This study is the first to report the presence of cytopathic virus in peripheral tissues following BCRV inoculation, with cytopathic virus being detected in the spleen and skeletal muscle for both BCRV-A– and BCRV-B–inoculated birds. This indicates that viral replication was actively occurring in peripheral tissues during the 3-day experiment and viral dissemination patterns of the two BCRV lineages were very similar. Although viremia was not detected in these birds, this is similar to previous studies of BCRV infections in birds that found low or no levels of viremia or oral shedding in birds after BCRV inoculation (Huyvaert et al. 2008, Brault et al. 2009).
The presence of viral RNA in blood and peripheral tissues following BCRV inoculation in this experiment is also in agreement with previous alphavirus studies in birds. Reisen et al. (2003) detected WEEV RNA in 5% of infected birds, 6 weeks postinoculation, whereas Huyvaert et al. (2008) detected BCRV RNA in the blood of 80% of birds 1–15 DPI, and BCRV RNA in the brain and skin of 50% of birds at 15 DPI. In this study, a high incidence of viral RNA in tissues was detected by both RT-PCR and a bead-based gene expression array. The higher frequency of viral RNA detection found in this study compared to previous avian alphavirus infection studies can likely be contributed to the time period of tissue collection. The samples in this study were collected during the acute infection stage (1–3 DPI) when viral dissemination into peripheral tissues is high (Ryman et al. 2007), compared to sampling during a later stage of infection that was measured in other studies (Reisen et al. 2003, Huyvaert et al. 2008).
Gene expression
In this study, both IFN-γ and IFN-α were significantly elevated in multiple tissues. Although the role of interferons in avian alphavirus infections is unknown, interferons are essential in the control of many other avian viruses, including avian influenza (Meng et al. 2011) and infectious bronchitis virus (Pei et al. 2001). Mildly elevated levels of type I and type II interferons are protective against VEEV and SINV infections in mammals (Grieder and Vogel 1999, Ryman et al. 2000) and likely contributed to the limited viral replication detected in this study.
Elevated levels of three PRRs (TLR7, TLR15, and RIG-1) were also detected in multiple tissue types following BCRV infection. In birds, TLR7 upregulation leads to an increase in proinflammatory cytokine release following infection with Marek's disease (Abdul-Careem et al. 2009) and avian influenza virus H9N2 infection (Xing et al. 2008). Additionally, TLR7 is protective against Ross River virus (an arthralgia-causing alphavirus in humans; Neighbours et al. 2012), and our results suggest that TLR7 is also important in the avian immune response to BCRV infections.
TLR15 has been identified in birds and reptiles, with yeast components as the only known agonists (Boyd et al. 2012). However, there may be a role for TLR15 in viral infections, because TLR15 is upregulated in the lung and intestine of chickens infected with avian influenza H9N2 (Nang et al. 2011). Upregulation of TLR15 in this study further supports the possibility of viral ligands for TLR15 and should be explored in greater depth.
RIG-1-like receptors are a class of cytoplasmic PRRs that contain RNA helicases that detect dsRNA and initiate antiviral signal transduction events. In birds, RIG-1 is required for a successful immune response against influenza virus (Barber et al. 2010). Although work on the effects of RIG-1 on alphavirus infections in vertebrates is inconclusive (White et al. 2011), the elevated RIG-1 levels reported here suggest that RIG-1 is also important in controlling avian BCRV infections.
In this study, TLR2, TLR3, TLR4, and CCR5 were elevated only in the skeletal muscle, which also had the highest viral RNA content of any tissue type (data not shown). The upregulation of additional genes in the skeletal muscle is likely related to the increased viral load and immune activation in this tissue. TLR2, TLR3, and TLR4 are upregulated in chickens following infection with infectious bronchitis virus (Wang et al. 2006, Guo et al. 2008), Marek's disease virus (Abdul-Careem et al. 2009), and avian influenza infection (Esnault et al. 2011). CCR5 is a chemokine receptor involved in leukocyte trafficking and is protective against neuroinvasive WNV disease in mice (Glass et al. 2005). However, the role of TLR2, TLR3, TLR4, and CCR5 in alphaviral infections is largely unknown. Taken together, the upregulation of TLR2, TLR3, TLR4 and CCR5 represent viral-specific immune responses that were likely protective in controlling viral replication in the skeletal muscle of BCRV-infected birds.
Conclusions
Previous studies of avian arbovirus infections have been mostly limited to virological and serological studies (McLean et al. 1995, Reisen et al. 2003, Reisen et al. 2006, Unnasch et al. 2006, Huyvaert et al. 2008) and descriptions of pathological damage of field-caught birds (O'Brien et al. 2010). This study is the first to report an avian immune gene expression profile following an alphavirus infection. Multiple gene sequences for a non-model bird (House Sparrow) were obtained, and a gene expression profile was produced using a multiplex bDNA expression assay system. The gene expression profile obtained is in agreement with work on alphaviruses in mammals and viral infections in birds in which multiple PRRs and type I and II interferons are produced that aid in the control of viral replication. In this study, the PRRs TLR7, TLR15, and RIG-1 were most ubiquitously upregulated, as were IFN-α and IFN-γ. A future comparison of the immune responses of susceptible and resistant birds to BCRV will deepen our understanding of the role of these genes in avian alphavirus infections.
Footnotes
Acknowledgments
We thank Emily Altrichter, Neil Backer, Sally Breining, Molly Hiatt, Tiffany Tran, and Charles Brown for their assistance. Funding was provided by a Creighton Summer Faculty Research Fellowship to Carol Fassbinder-Orth and a Creighton Ferlic Summer Fellowship to Virginia Barak. The project described was supported by grant no. G20RR024001from the National Center for Research Resources. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Center for Research Resources of the National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist.