
Abstract
Human encroachment on wildlife habitats has contributed to the emergence of several zoonoses. Pathogenic hantaviruses are hosted by rodents and cause severe diseases in the Americas and Eurasia. We reviewed several factors that potentially drive prevalence (the proportion of infected rodents) in host populations. These include demography, behavior, host density, small mammal diversity, predation, and habitat and landscape characteristics. This review is the first to include a quantitative summary of the literature investigating hantavirus prevalence in rodents. Demographic structure and density were investigated the most and predation the least. Reported effects of demographic structure and small mammal diversity were consistent, whereby reproductive males were most likely to be infected and prevalence decreased with small mammal diversity. The influences of habitat and landscape properties are often complex and indirect. The relationship between density and prevalence merits more investigation. Most hantavirus hosts are habitat generalists and their control is challenging. Incorporating all potential factors and their interactions is essential to understanding and controlling infection in host populations.
Introduction
I
Hantaviruses (family Bunyaviridae, genus Hantavirus) are rodent-borne RNA viruses (Lee et al. 1978, Schmaljohn et al. 1985, Vaheri et al. 2013) and etiologic agents of hemorrhagic fever with renal syndrome (HFRS) and hantavirus pulmonary syndrome (HPS) in Eurasia and the Americas, respectively (Nichol et al. 1993, Vapalahti et al. 2003). Hantaviruses are considered to be host specific (Vapalahti et al. 2003), yet there is spillover to other species of rodents and shrews (Schmaljohn and Hjelle 1997, Klingström et al. 2002, Schmidt-Chanasit et al. 2010). Shrews, moles (order Eulipotyphla), and bats (order Chiroptera) can be genuine hosts to several hantavirus species (Gu et al. 2013), but only rodents (families Cricetidae and Muridae) have so far been found to host disease-causing hantaviruses (Lee et al. 1978, Nichol et al. 1993, Olsson et al. 2010).
Hantaviruses are a global health concern, with approximately 150,000–200,000 human disease cases occurring yearly worldwide (Schmaljohn and Hjelle 1997, Bi et al. 2008). Human infections mostly result from inhalation of aerosolized viral particles excreted or secreted by infected rodents (in, urine, feces, and saliva) into the environment. In the Americas, the mortality rate due to HPS is about 35% (MacNeil et al. 2011), whereas in Eurasia, HFRS is less severe, with a mortality rate of 12% or less depending on the disease-causing virus species (Bi et al. 2008). However, HFRS causes considerable suffering in patients, especially when complications occur (Niklasson 1992, Vaheri et al. 2013).
Hantaviruses are transmitted horizontally within host populations (Yanagihara et al. 1985), either directly through aggressive interactions and grooming, or indirectly through the environment. In the latter scenario, infection occurs when susceptible individuals inhale infectious particles shed into the environment (Hardestam et al. 2008). As host populations increase in density and distribution, the likelihood for contact with humans increases (Niklasson and LeDuc 1987, Nicklasson et al. 1995, Mills et al. 1999, Yates et al. 2002, Olsson et al. 2003, Pettersson et al. 2008, Kallio et al. 2009, Olsson et al. 2009). Both the number of infected rodents, related to their density and virus prevalence (the proportion of infected rodents in the host population), and abiotic properties of the environment determine the environmental viral load and thus human risk of acquiring infection (Reusken and Heyman 2013).
Within the hantavirus transmission cascade, the distribution and prevalence of infection in rodent hosts is influenced by a range of factors (Fig. 1). They often interact or work in accordance or opposition to give temporally and spatially variable outcomes (Mills et al. 2010). The multifactorial nature of the hantavirus infection system (Reusken and Heyman 2013) is complicated by factors that are both direct and indirect modifiers of prevalence in populations. For example, small mammal species diversity (from here on referred to as biodiversity) may modify density and behavior of host populations (Suzán et al. 2009, Carver et al 2011a). A decrease in host density or intraspecific encounter rates may lower prevalence, but through distinct pathways. Moreover, biodiversity and rodent host density are affected by habitat suitability and quality (Ecke et al. 2002).
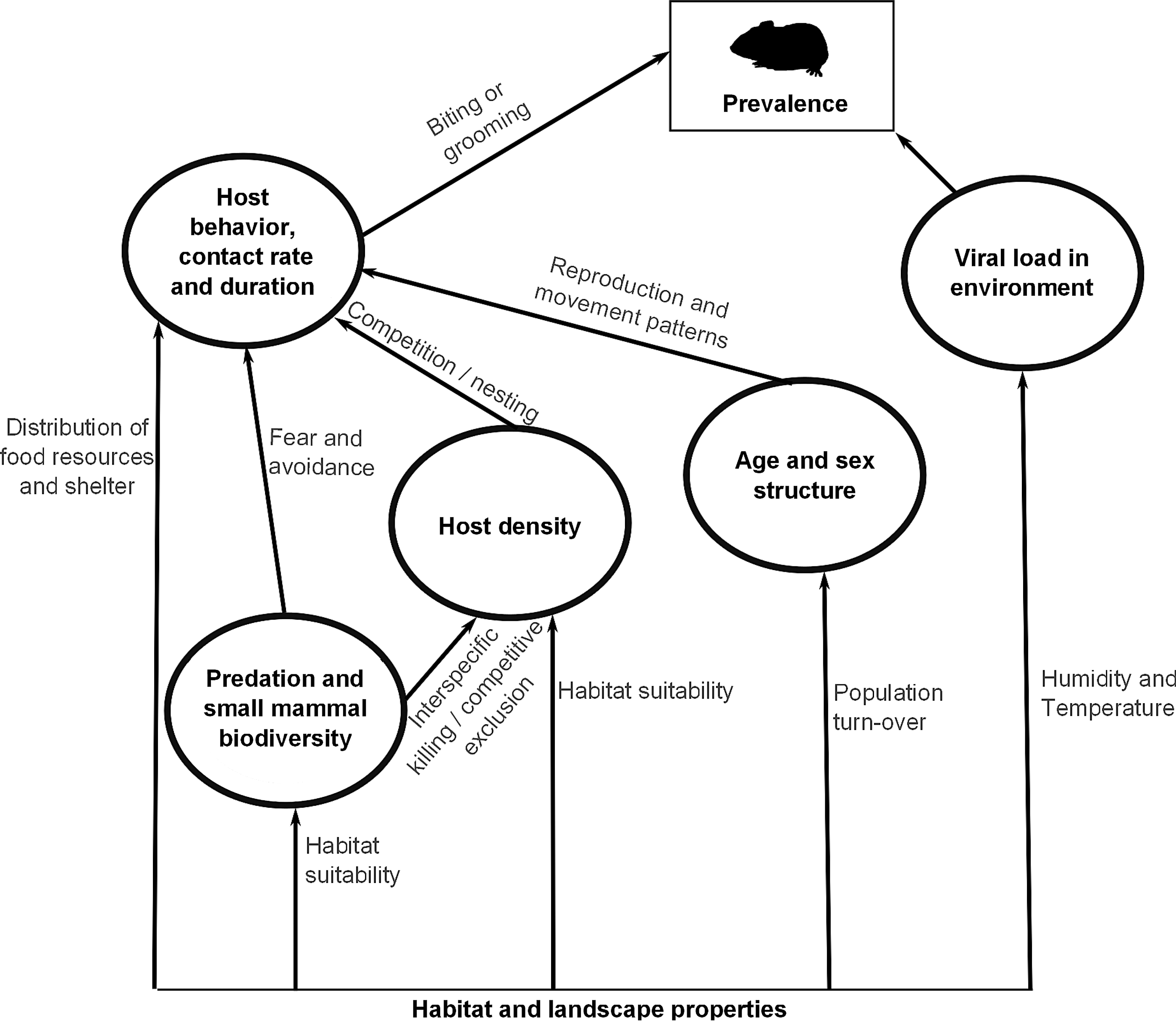
The relationship among different explanatory factors and prevalence. The infection system consists of several interacting factors through which prevalence is directly and indirectly modified.
Understanding what drives infections in host populations can help establish an early warning system to minimize human incidence and take measures to reduce infection in hosts (Mills 2005, Chomel et al. 2007, Olsson et al. 2009). A large body of literature has discussed the ecology of hantaviruses and their reservoirs (for reviews, see Mills et al. 1999, Mills 2005, Dearing and Dizney 2010, Mills et al. 2010, Olsson et al. 2010) and attempted to identify endemic areas and high-risk habitats (Yahnke et al. 2001, Olsson et al. 2005, Salvador et al. 2011, Voutilainen et al. 2012).
We are aware of the importance of strictly indirect drivers of hantavirus prevalence, such as factors involved in a trophic cascade leading to higher numbers of infectious rodents (Yates et al. 2002). Regardless of the relationship between host density and prevalence, the number of infected rodents may still increase with host density (Davis et al. 2005). Indeed, understanding the relationship between densities and phenomena such as mast years (years with high tree seed production) (Tersago et al. 2009, Clement et al. 2009), El Niño (Yates et al. 2002), and rodent cycles (Hörnfeldt 1994, Saitoh et al. 1998, Olsson et al. 2002) aids in assessing disease outbreak risk (see Linard et al. 2007).
Our aim here is to give a quantitative summary of the literature pertaining to factors driving prevalence (see above) in host populations. This review will be the first to focus on prevalence and to identify, by numbers, where the bulk of scientific attention and evidence lies and what factors affected hantavirus prevalence consistently or idiosyncratically among studies. We also highlight understudied factors, thus revealing gaps in scientific knowledge and paving the way for future investigations. Upon surveying the literature, we identified the following factors as potentially modifying prevalence in rodent host populations through at least one direct route: (1) Intrinsic host population factors, including behavior, demography, density, and survival of infected rodents, and (2) extrinsic factors, such as habitat and landscape, biodiversity, and predation.
We follow the schematic presentation of the infection system (Fig. 1). Thus, our discussion begins with intrinsic population properties and ends with extrinsic factors. We will qualitatively address interactions among factors in the discussion.
Methods
We performed the literature search using Web of Science and included studies published from as early as possible and through June, 2013. We chose key terms based on the potential intrinsic and extrinsic factors listed in the introduction plus three more general terms to broaden our search and retrieve studies that failed to appear in specific search queries. We used the following key terms with the three general ones coming first: 1. Hantavirus ecology 2. Hantavirus dynamics 3. Hantavirus prevalence 4. Hantavirus density dependence and hantavirus prevalence abundance 5. Hantavirus landscape 6. Hantavirus habitat 7. Hantavirus biodiversity 8. Hantavirus predator and hantavirus predation 9. Hantavirus demography and hantavirus age sex 10. Hantavirus behavior (behavior) 11. Hantavirus survival and hantavirus fitness
We only included articles published in peer-reviewed journals and excluded articles strictly based on mathematical models because our aim was to summarize empirical findings. We also excluded articles not related to hantavirus prevalence in rodent hosts, such as those addressing physiological aspects of hantaviral infections or infection patterns in humans. Most studies considered the effects of more than one factor, which may render classifications in our summary interdependent. However, detecting an effect of a single factor necessitates accounting and controlling for contextual or confounding ones.
Results and Discussion
Using the listed search terms, we retrieved 86 unique articles (see Appendix 1). Most studies, probably due to the high fatality of HPS, are from the Americas (Fig. 2). Overall, the relationship between demographic factors (age and sex) and prevalence was investigated the most (age, 44 articles; sex, 48 articles) (Fig. 3). Conversely, influences of infection status and fitness (15), biodiversity (13), behavior, wounds and scars (13), movement (4), landscape (6), and predation (1) were examined less. The relationship between demographic factors and prevalence has been considered from 1989 onward, whereas that between biodiversity and prevalence has been highlighted more recently (Fig. 4). In the following sections, we summarize and discuss different factors in light of their standing in the literature, potential and reported interactions, and future perspectives.
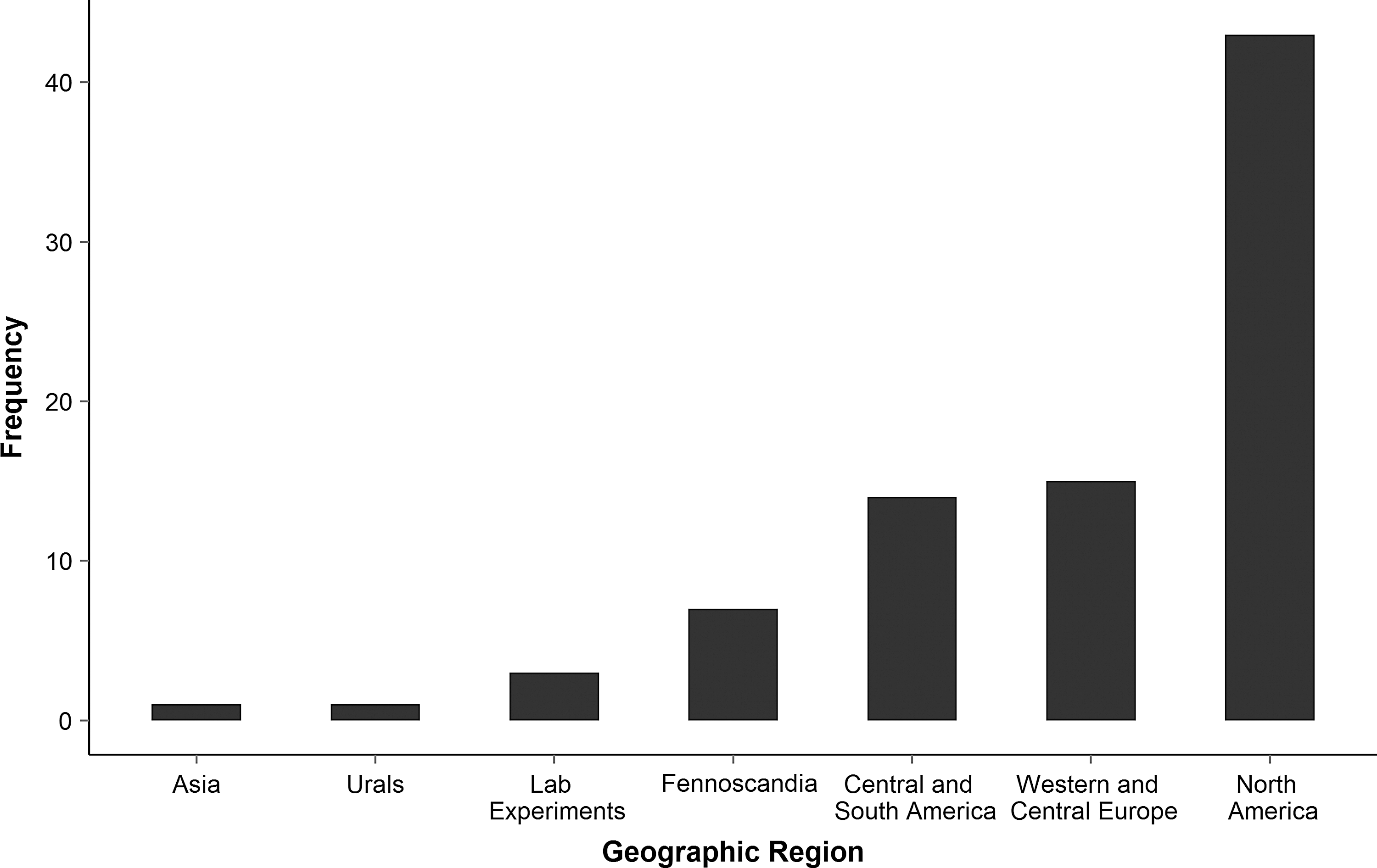
Geographic distribution of hantavirus prevalence studies. Most studies of prevalence in hosts come from the Americas, where mortality from hantavirus pulmonary syndrome is about 35%. Studies on the bank vole–Puumala hantavirus system are common and were conducted in Fennoscandia and western Europe.
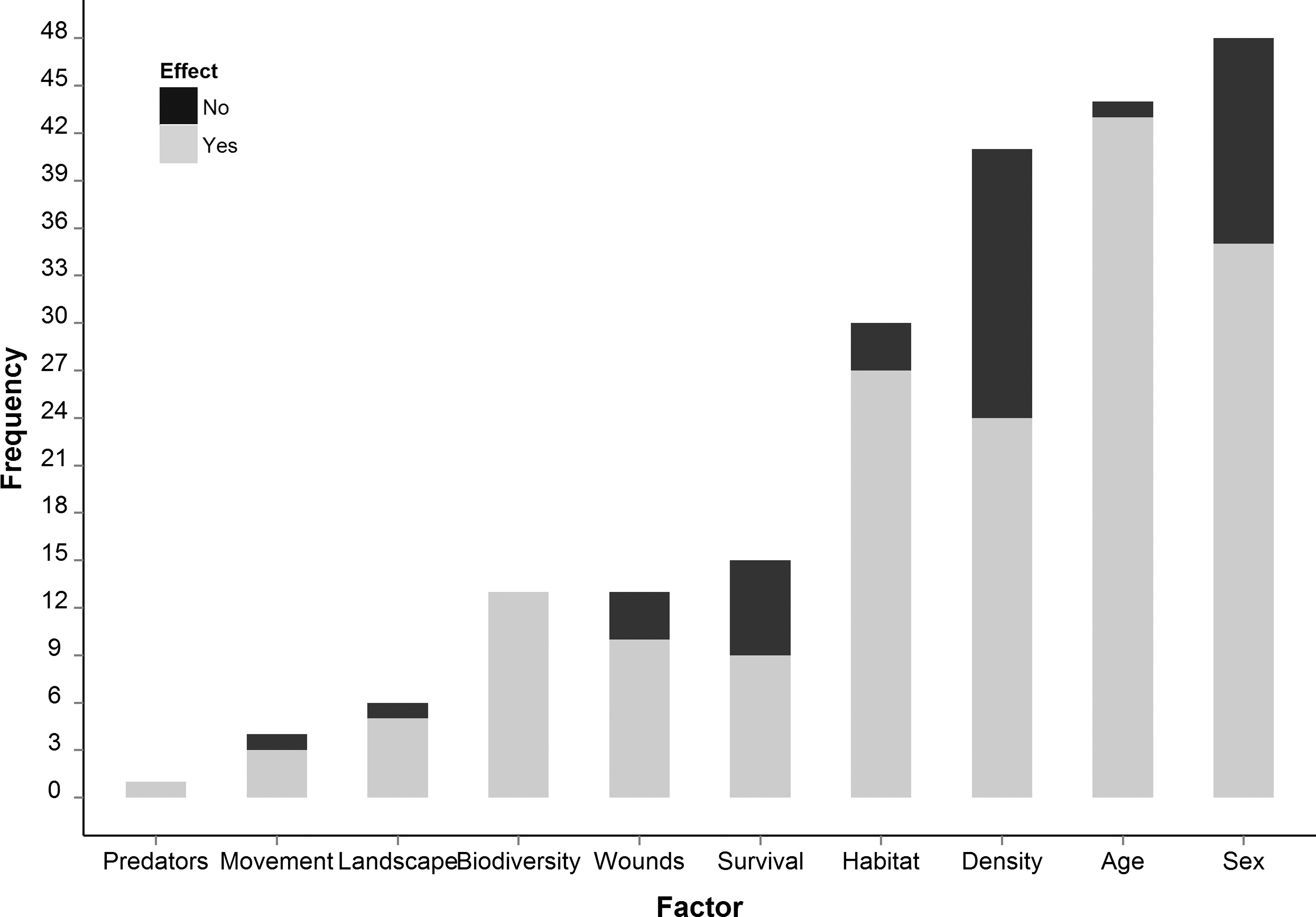
Quantitative summary of the literature in relation to factors modifying prevalence. Demography (age and sex) is the factor considered most in the literature. Landscape characteristics, behavior (wounds/scars and movement), biodiversity, and predation have been less studied in relation to prevalence. The relationship between density and prevalence is inconsistent in the literature, as only 59% of studies found a relationship.
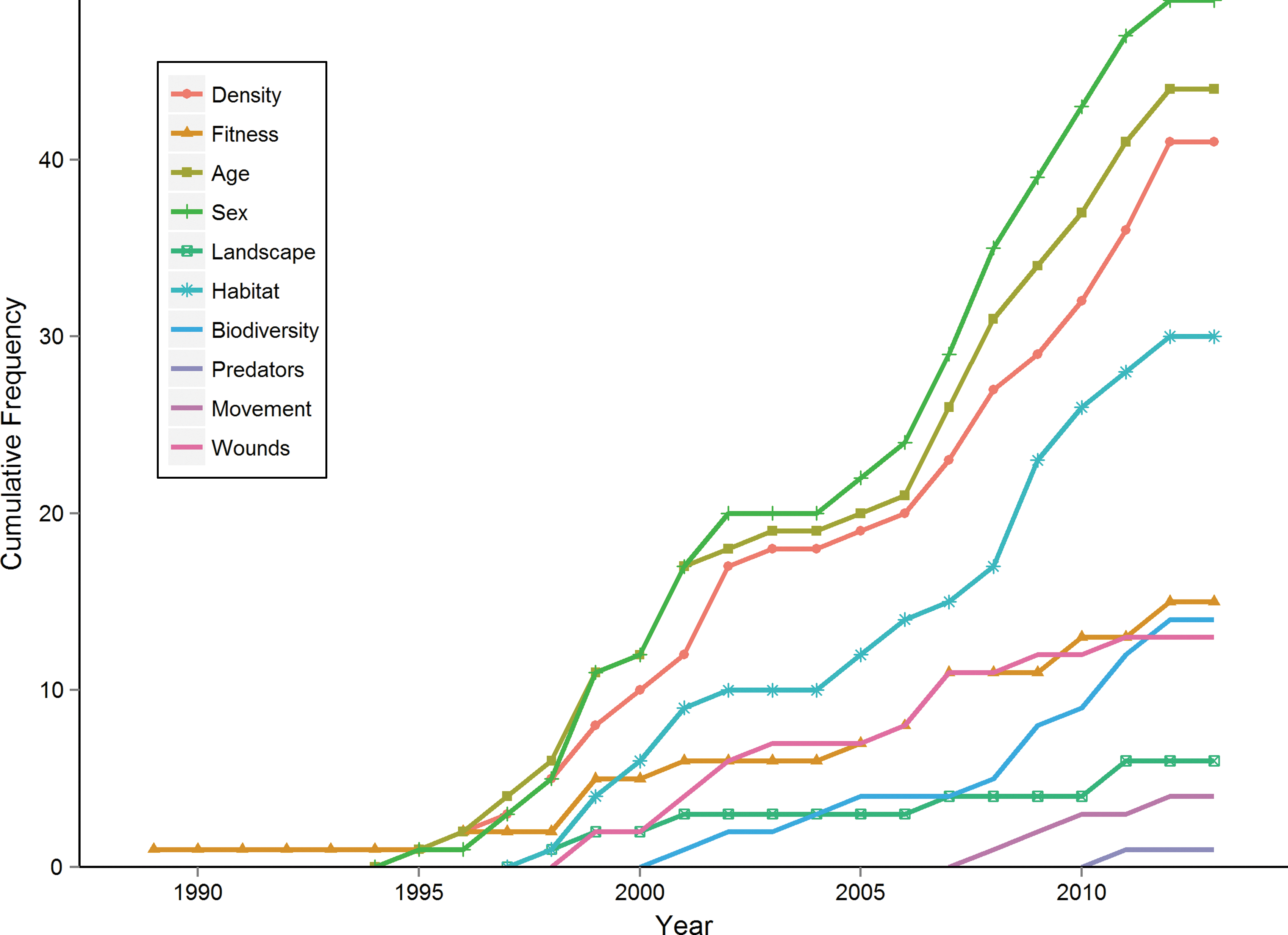
The cumulative temporal trend of studies investigating factors driving hantavirus prevalence in rodent hosts that are reviewed here. Recently, studies have investigated the relationship between biodiversity and landscape properties and prevalence.
Demography, behavior, and prevalence
Infection probability in hosts increases with age and is higher in males. Forty-three out of 44 studies found an effect of age and 35 out of 48 found males more likely to be infected than females. Results were consistent across different hantavirus–host systems; for example: bank vole, Puumala hantavirus in the Urals (Bernstein et al. 1999); bank vole, Puumala hantavirus in Sweden (Olsson et al. 2002); Vesper mouse (Calomys laucha), Laguna Negra hantavirus in Paraguay (Yahnke et al. 2001); deer mice (Peromyscus maniculatus), Sin Nombre hantavirus in North America (Kuenzi et al. 2001).
Hantaviruses are transmitted horizontally in their host population, and age- and sex-specific behaviors may affect infection probability (Mills et al. 1999). Most studies suggest that during the breeding season the virus is primarily transmitted through intraspecific aggressive behaviors such as biting (Root et al. 1999, Pearce-Duvet et al. 2006, Calisher et al. 2007). Ten out of 13 studies found that wounds and scars are related to antibody status. Tersago et al. (2011) reported that sex differences in infection status exist in the reproductive portion of the population. Outside the breeding season, adults did not suffer higher infection risk. This suggests a seasonal variation in risk behavior because communal nesting in winter may promote transmission directly through grooming and indirectly through the environment (Calisher et al. 1999).
Although there is a relationship between aggression and infection status, some studies have suggested that this relationship is complex (Mills 2005) and might result from physiological modifications caused by the infection itself. For example, Douglass et al. (2007) reported that acquisition of scars was higher in the breeding season, but found no relationship between scar acquisition and acquiring infection. They hypothesized that infection status may affect behavior and promote aggression. In a laboratory study, Klein et al. (2004) had earlier found that infected male rats (Rattus norvegicus) were more likely to initiate aggressive interactions and spent longer periods engaged in aggressive behaviors compared to noninfected males.
Another potential risk factor associated with age is the loss of maternal antibodies. Litters of infected dams are immune for approximately 3 months (Kallio et al. 2006b). Thus, individuals become susceptible after the loss of maternal antibodies as subadults. The increasing risk with age is exacerbated by male dispersal, during which dispersers may encounter and engage in aggressive behavior with potentially infected individuals (Dobly et al. 2012).
Future demographic studies should aim at establishing causal mechanisms linking behavior, demography, and infection status. Controlling for the association between age and wounds and scars (Escutenaire et al. 2002) is necessary to rule out age as a confounding factor and establish a genuine relationship between the reproductive functional group and infection risk.
Host density and prevalence
Transmission in horizontally transmitted viruses is commonly density dependent (Adler et al. 2008), and contact rates are expected to increase with host density, culminating in higher prevalence (Mills et al. 1999). However, the association between prevalence and host density in the literature is inconsistent. Out of 41 articles, only 24 (59%) found evidence for a relationship between density and prevalence, whereas 17 (41%) found no relationship. Even within the same hantavirus–host system, results differed. In North America, Boone et al. (2002) in a study from 1995 to 1998 found that, despite a decline in deer mice populations, prevalence did not decline, whereas in a longer 10-year study, Calisher et al. (2007) did find a positive relationship between density and prevalence.
There are several variables in an infection system that could modify or mediate the relationship between density and prevalence. The influx of a large number of uninfected newborns into a host population may temporarily mask the effects of increased contact rates as observed in studies finding a negative relationship between density and prevalence (Lehmer et al. 2012). Also, maternal antibodies result in a negative relationship between prevalence in the prebreeding season and prevalence at the end of it (Kallio et al. 2010), as influx of immune individuals delays virus spread in a population (Adler et al. 2008). Indeed, some studies have found evidence for delayed density dependence, whereby prevalence is correlated with past densities (Niklasson et al. 1995, Madhav et al. 2007).
An increase in prevalence is a consequence of increasing contact rates and transmission with density. Yet, as outlined below, contact rates are not determined solely by density, and using density as a surrogate for contact rates might be misleading. Differences in habitat characteristics may alter movement and/or contact rates. In habitats with a patchy distribution of resources, individuals might move longer distances in search of food and may have higher contact rates compared to individuals in a homogeneous habitat (Root et al. 1999). Consequently, even at comparable densities, habitats with different resource distribution, shelter availability, or structural composition may be characterized by different host contact rates. Thus, contact rates may not necessarily depend on density per se but on the per capita availability and spatial distribution of resources and shelter. Even in the case of a positive relationship between contact rate and density, contact rates may become saturated beyond a density threshold (Adler et al. 2008). And if the virus goes locally extinct below a certain host density (Boone et al. 1998), then prevalence will only be related to density between two threshold values—a lower bound that corresponds to the minimum number of hosts for virus maintenance in a population and an upper bound at which saturation in contact rates occurs.
Additionally, an increase in stress in host populations at high densities may increase susceptibility to disease (Mills 2005). Consequently, transmission within host populations would accelerate leading to a high prevalence at peak densities in affected functional groups. Furthermore, comparing prevalence across different spatial or temporal scales must take into account the demographic structure of populations. In North America, prevalence was found to be lower in the part of the study area with a high population turnover and higher proportion of younger individuals (Calisher et al. 2001). Demographic heterogeneity in host populations not only has implications on the differential likelihood of being infected, but also on transmission efficiency. Larger adults may be responsible for most of the contacts in a population (Clay et al. 2009a). Also, high population density might delay maturation of juveniles (Glorvigen et al. 2012). As a result, subadults remain in their natal nest and might acquire infection from their mother after losing their maternal antibodies. Tersago et al. (2012) suggested that hantavirus infection accelerates maturation in subadults, which would facilitate infection spread in a population due to behavioral correlates of reproductive activity (see previous section). Furthermore, higher density might increase the transmission rate through elevated viral load in the environment. Because virus persistence is mediated by humidity and temperature (Kallio et al. 2006a, Voutilainen et al. 2012), the effectiveness of this indirect transmission route likely differs across habitats, seasons, and years.
The multitude of factors operating on prevalence might easily mask the relationship between density and prevalence in host populations. To detect an effect of density, investigators ought to control for demographic, behavioral, habitat-related, and seasonal variations in host populations statistically and when designing experiments or sampling regimes. Long-term studies at a regional scale (see Niklasson et al. 1995, Calisher et al. 2007, Carver et al. 2011b) may be more appropriate to evaluate the effect of density on prevalence compared to studies conducted on a shorter time frame or those with a small sample size. Results of these latter studies may be inconclusive due to unreliable prevalence estimates and short-term variations.
Infected host survival and prevalence
Having co-evolved with a specific host over a long period, hantaviral infections in rodent reservoirs were considered asymptomatic (Childs et al. 1989, Bernstein et al. 1999). However, recent evidence suggests that hantaviral infections impair winter survival (Kallio et al. 2007), hinder weight gain in its host (Douglass et al. 2007), and increase juvenile mortality (Douglass et al. 2001). The maintained immune response to chronic hantaviral infection might be energetically costly and reduce investment in maintenance. Additionally, the development of diseases may be triggered by stressful conditions, such as crowding and unfavorable weather (Mills 2005, Niklasson et al. 2006 and references therein). If host survival is affected by hantavirus infection, then prevalence may thus vary, depending on the timing of population sampling. A lower over-winter survival of infected individuals compared to that of uninfected ones may modify short-term variation in prevalence and confound the influence of other drivers. Of 15 studies investigating the effects of hantavirus infection on host longevity, seven found negative effects, whereas two found a positive relationship between infection and longevity and six found no relationship. However, to our knowledge, no study has yet considered the impact of differential survival within host populations on short-term variation in prevalence.
The positive relationship between longevity and infection status in two studies (Abbot et al. 1999, Owen et al. 2010) could be due to the confounding effect of age and status. As older established residents have a higher chance of surviving adverse conditions (Abbot et al. 1999) and a higher likelihood of being infected, the relationship between longevity and infection status is unlikely to be genuine. Ecological studies may be unable to establish a causal association between infection status and fitness due to the multitude of co-varying or confounding elements. Laboratory approaches may be better suited to disentangle the consequences of age and infection.
Biodiversity and prevalence: The dilution effect
The negative association between biodiversity and infection prevalence is consistent across studies and hantavirus–host systems. All of the 13 studies reported a negative relationship between biodiversity and prevalence, two of which involved experimental manipulation (Clay et al. 2009b, Suzán et al. 2009). Two nonmutually exclusive mechanisms may give rise to this relationship (Keesing et al. 2006, Clay et al. 2009b). The presence of nonhost rodent species may: (1) indirectly lower prevalence by suppressing host density (susceptible host regulation) or (2) directly lower prevalence through reducing contact rate and duration (encounter reduction). A reduction in host density or alteration of size and configuration of host territory is expected to reduce prevalence through density-dependent mechanisms. When nonhost rodents are present, the proportion of interactions among rodents that result in transmission events is expected to decrease. Even if there is an infection spillover, infected nonhost individuals will function as viral sinks and a dead end in the transmission cycle. This pathway acts on prevalence indirectly through the density-dependent pathway discussed above and is subject to the same conditions and caveats. Nevertheless, the presence of nonhost rodent species may modify prevalence directly through reducing duration and number of host intraspecific interactions. This may be especially important because hantaviruses are transmitted less efficiently than other viruses (Botten et al. 2002). Hence, even if host density is unaffected, the presence of competing species may lower prevalence (Clay et al. 2009b).
Biodiversity depends amongst others on habitat suitability (Yahnke et al. 2001). Anthropogenic disturbance may result in local extinction of specialist species of small mammals. This releases generalist species, including hantavirus hosts, from interference competition, culminating in an increase in their density. An increase in density of hosts may lead to an increase in prevalence (see section on density and prevalence) and human risk of infection (Mills 2006). However, it is challenging to separate the effects of habitat and landscape (see next section; Fig. 3) from those of biodiversity on host density, demography, behavior, and ultimately prevalence. Studies involving experimental manipulation that control for factors such as habitat (see Suzán et al. 2009) and host density (see Clay et al. 2009b) may better highlight the influence of biodiversity on hantavirus transmission and prevalence. Moreover, temporal variation in biodiversity ought to be accounted for (e.g., Carver et al. 2011a). Failing to do so may mask the effect of both biodiversity and density on prevalence.
Habitat, landscape characteristics, and prevalence
Of 30 studies investigating the effect of habitat on prevalence, 27 found an association. Landscape properties were considered in six studies and were found to be associated with prevalence in five cases. Habitat and landscape properties affect biodiversity (Lehmer et al. 2012), host density (McIntyre et al. 2005), host behavior (Root et al. 1999), and demography (Calisher et al. 2001). The influence of habitat and landscape characteristics on infection prevalence is complex. Often, habitat and landscape characteristics modify prevalence indirectly through various ecological factors both intrinsic and extrinsic to host populations (Linard et al. 2007). Landscape characteristics such as connectivity and fragmentation are related to prevalence indirectly through their effect on movement, dispersal, and survival in host populations (Langlois et al. 2001). The direct route through which habitat characteristics may affect prevalence is proposed to be through modifying virus survival and thus viral load in the environment (Voutilainen et al. 2012).
Because each rodent host species is associated with a unique suite of rodent competitors, predators, and habitat preferences, there is perhaps no universal habitat or landscape effect. In subsequent paragraphs we briefly discuss some facets of the direct and indirect relationship between prevalence and habitat and landscape properties.
High-quality habitats sustain larger host densities, which may lead to increased prevalence through direct or delayed density-dependent mechanisms (McIntyre et al. 2005). Moreover, individuals dispersing through or residing in lower-quality or disturbed habitats may suffer increased mortality due to resource scarcity or predation. This leads to a fast population turnover and skews the population structure toward younger individuals (Lehmer et al. 2008). In contrast, higher-quality habitats may support heavier and older resident males that are more likely to be infected (Calisher et al. 2001, Goodin et al. 2009). Abbot et al. (1999) reported that during adverse conditions, suitable habitat patches are able to sustain focal host populations from which the virus spreads during subsequent recolonization of marginal habitats. In marginal patches, low host density may not be sufficient to sustain the virus (Guivier et al. 2011).
Other studies found prevalence to be higher in low-quality habitats and proposed the following explanation. Although higher-quality habitats sustain higher densities of rodent host, they still provide more per capita nesting sites and resources compared to lower-quality habitats. Hence, host aggregation and intraspecific encounters are expected to be less in high-quality habitats (Pearce-Duvet et al. 2006). Heterogeneous resource distribution in low-quality habitats increases host movement, competition, and thus risk of infection (Root et al. 1999, Heyman et al. 2009). Langlois et al. (2001) also found prevalence to be higher in more isolated, lower-quality patches.
Human-induced habitat alterations will invariably be related to quality, but the response of the small mammal community in a given habitat to anthropogenic disturbance is species specific (Yahnke et al. 2001, Mills 2005). Most hantavirus hosts are habitat generalists and dominate disturbed areas (Ruedas et al. 2004, Mills 2005, Mills 2006). In Central and South America, for example, agricultural fields could sustain higher host density due to higher food provisioning and releasing hosts from interspecific competition and predation (Yahnke et al. 2001). Thus, through the biodiversity route, anthropogenic disturbance may lead to higher prevalence. However, there is no a priori reason for predicting a general disturbance effect. In Utah in the United States, disturbance from use of all-terrain vehicles led to higher prevalence (Mackelprang et al. 2001). As vehicles damaged the vegetation, altered soil properties, and increased fragmentation, suitable patches became smaller with higher density of deer mice, the hosts of the Sin Nombre virus. The authors suggested that through density-dependent and behavioral mechanisms, prevalence in habitats disturbed by motor vehicles increased (Mackelprang et al. 2001).
Indirect transmission through the environment is necessary for the long-term persistence of the virus in a population (Sauvage et al. 2003). Virus survival is proposed to be driven by moisture and temperature (Kallio et al. 2006a). Forests with dense canopy have lower forest floor temperature, higher humidity, and get less ultraviolet (UV) light penetration (Goodin et al. 2009). Consequently, these environments ought to have a higher viral load compared to forests with a sparser canopy. Sauvage et al. (2002) reported that prevalence was higher in traps placed in north-facing slopes and suggested higher humidity as the explanation. All else being equal, it is intuitive that prevalence would increase with viral load in the environment (Voutilainen et al. 2012). However, according to our knowledge, no field study has directly studied the relationship between abiotic habitat properties and prevalence.
Microhabitat structure, patch isolation, and resource distribution in a given habitat may dictate host movement patterns and contact rates. These factors have not been frequently investigated (but see Langlois et al. 2001, Guivier et al. 2011) and ought to be considered in combination with density to better understand virus transmission and prevalence. Also, hantavirus–host systems are not uniform and may be driven by different factors in different biomes and geographic regions. Determinants of hantavirus dynamics could thus be different in the boreal zone compared to the subtropics (Owen et al. 2010, Olsson et al. 2010).
Predation and prevalence
Although their precise role in regulating rodent population densities and multiannual cycles is still debated, predators affect rodent populations (Hanski et al. 2001, Korpimäki et al. 2005). Surprisingly, yet perhaps due to lack in data or resource, their role in hantavirus ecology has not been investigated. Only one study considered the effects of predator richness on hantavirus prevalence (Orrock et al. 2011). The authors found that rodent host populations in islands with higher predator species richness exhibited lower prevalence compared to predator poor ones.
Predators may regulate prevalence in rodent hosts in several ways (for review, see Ostfeld and Holt 2004). If predators are more likely to select an infected prey, then predation reduces prevalence directly (Packer et al. 2003). Infection may force infected individuals to increase their foraging activity due to energetic costs of maintained immune response, rendering them more susceptible to predation. Additionally, age- and sex-specific behaviors that increase the risk of hantavirus exposure may also increase predation risk. Male rodents, especially those weighing less, were overrepresented in the diet of avian predators (Koivunen et al. 1996), owing perhaps to higher exposure when dispersing or when forced out of high-quality habitats by established resident males. A male bias in rodents in the diet of predators may alter the sex structure of the host population. This leads to a reduction of both the number of individuals at higher risk of hantavirus exposure and infected individuals already involved in virus transmission.
Through instilling fear and avoidance behaviors, the presence of predators may limit movement and contact rates within a host population (Brown et al. 1988). In forest patches with dense canopy cover, avian predation risk may be lower, thus leading to increased host movement and survival (Polop et al. 2010). Consequently, higher prevalence in such patches could be related to both longer persistence of the virus in the environment and reduced predation risk. However, this latter pathway has not yet been studied.
Even without selective predation, predators may reduce prevalence by suppressing densities of host populations (Ostfeld and Keesing 2012). This indirect route nevertheless depends on the relationship between density and prevalence discussed earlier.
Conclusions
Aside from exposing the complexity of hantavirus–host systems, our review quantitatively classified factors in relation to their reported effects on prevalence. Infection risk in hosts is higher in reproductive males compared to females and nonreproductives. The spread of infection is hence associated with the reproductive functional group. Moreover, biodiversity dilutes infection both directly through reducing host density and indirectly through altering its behavior.
The role of habitat and landscape properties is context dependent and mostly related to modification of other factors, such as host demography, host density, biodiversity, and viral load in the environment. However, the evidence for a relationship between density and prevalence remains inconclusive.
Certain aspects of the infection system have been considered less than others in the literature (e.g., predation and landscape characteristics) because their investigation is more resource demanding. Nevertheless, it remains relevant to consider predation as a potential modifier of prevalence because it may suppress host density, selectively remove infected individuals or those at a higher risk of infection, and modify host behavior. With increasing human encroachment on wildlife habitats, there is an increased risk of emergence and re-emergence of zoonoses. Hantavirus hosts occupy large geographic ranges and have high intrinsic population growth rates, and controlling their numbers may be challenging. To forecast human exposure risk, we can monitor ecological variables within the trophic cascade that culminates in high host densities. However, a multidisciplinary endeavor that incorporates all potential factors and their interactions is essential to implement proactive rather than reactive measures when designing local- and regional-scale strategies for monitoring and controlling infection in host populations.
Footnotes
Author Disclosure Statement
No competing financial interests exist