
Abstract
Vector-borne disease (VBD) pathogens remain an emerging health concern for animals and humans throughout the world. Surveillance studies of ticks and humans have made substantial contributions to our knowledge of VBD epidemiology trends, but long-term VBD surveillance data of dogs in the United States is limited. This seroreactivity study assessed US temporal and regional trends and co-exposures to Anaplasma, Babesia, Bartonella, Borrelia burgdorferi, Dirofilaria immitis, Ehrlichia spp., and spotted fever group Rickettsia in dogs from 2004–2010. Dog serum samples (N=14,496) were submitted to the North Carolina State University, College of Veterinary Medicine, Vector Borne Disease Diagnostic Laboratory for vector-borne pathogens diagnostic testing using immunofluorescent antibody (IFA) and enzyme-linked immunosorbent assay (ELISA) assays. These convenience samples were retrospectively reviewed and analyzed. The largest proportion of samples originated from the South (47.6%), with the highest percent of seroreactive samples observed in the Midatlantic (43.4%), compared to other US regions. The overall seroreactivity of evaluated VBD antigens were Rickettsia rickettsia (10.4%), B. burgdorferi (5.2%), Ehrlichia spp. (4.3%), Bartonella henselae (3.8%), Anaplasma spp. (1.9%), Bartonella vinsonii subsp. berkhoffii (1.5%), Babesia canis (1.1%), and D. immitis (0.8%). Significant regional and annual seroreactivity variation was observed with B. burgdorferi, Ehrlichia, and Rickettsia exposures. Seasonal seroreactivity variation was evident with Rickettsia. Seroreactivity to more than one antigen was present in 16.5% of exposed dogs. Nationally, the most prevalent co-exposure was Rickettsia with Ehrlichia spp. (5.3%), and the highest odds of co-exposure was associated with Anaplasma spp. and B. burgdorferi (odds ratio=6.6; 95% confidence interval 5.0, 8.8). Notable annual and regional seroreactivity variation was observed with certain pathogens over 7 years of study, suggesting canine surveillance studies may have value in contributing to future VBD knowledge.
Introduction
V
With similar environmental and vector exposures as their human counterparts, dogs can be effective sentinels for human exposure to VBDs (Nicholson et al. 2010, Mead et al. 2011). Depending upon the VBD pathogen(s), clinical and pathologic abnormalities can induce highly variable clinical presentations and disease severity (Nicholson et al. 2010, Chomel 2011). Dogs and humans infected with VBD pathogens can exhibit similar symptoms, with typical clinical signs including fever, lethargy, anorexia, lameness, myalgia, and thrombocytopenia (Bowman et al. 2009, Nicholson et al. 2010). Improved understanding of the geographic distribution of VBD pathogens can facilitate prompt disease diagnosis and effective animal and public health interventions.
A diverse spectrum of vectors is associated with VBD pathogens, and vector ecology contributes to pathogen distribution and disease occurrence. New vectors continue to be identified for both established and emerging VBD pathogens. For example, Dermacentor variabilis and Dermacentor andersoni are established vectors for Rickettsia rickettsii, the pathogen associated with Rocky Mountain spotted fever (RMSF) (McQuiston et al. 2011, Allison and Little 2013). New tick species, such as Rhipicephalus sanguineus in the Southwest, and Amblyomma americanum in the Midwest, have been recently associated with R. rickettsii exposure in the United States (Diniz et al. 2010, Openshaw et al. 2010, Berrada et al. 2011, Allison and Little 2013). Other spotted fever group (SFG) rickettsiae, such as R. parkeri and R. amblyommii, have been identified in multiple tick species in the United States, and R. felis has been identified in fleas (Grasperge et al. 2012, Allison and Little 2013). Arthropods are common vectors for many canine VBDs, including the mosquito vector of Dirofilaria immitis, and the flea vector (and a potential tick vector) for Bartonella spp. (Diniz et al. 2010, Chomel 2011).
Both vector and host species can harbor multiple pathogens, creating potential for complex exposures and concurrent co-infections with more than one VBD pathogen. Co-infections with VBD pathogens pose diagnostic and treatment challenges, including atypical clinical presentations, increased illness severity, and incomplete treatment response in domestic animals and humans (Beall et al. 2008, Gaunt et al. 2010, De Tommasi et al. 2013). The epidemiology and pathogenesis of tick borne co-infections have not been clearly elucidated, and the reported prevalence of co-exposures has varied widely in tick, human, and domestic animal studies (Swanson et al. 2006, Beall et al. 2008, Bowman et al. 2009, Nieto and Foley 2009, Otranto et al. 2009, Beall et al. 2012). Using a database of samples submitted by veterinary clinicians from dogs suspected of a VBD infection, we retrospectively assessed US seroreactivity temporal and regional trends and co-exposures of Anaplasma, Babesia canis, Bartonella spp., Borrelia burgdorferi, D. immitis, Ehrlichia, and Rickettsia spp. from 2004–2010.
Materials and Methods
Study population
Canine serum samples submitted to the North Carolina State University, College of Veterinary Medicine, Vector Borne Disease Diagnostic Laboratory (VBDDL) between January 1, 2004, and December 31, 2010, were selected for study. Samples originated from US veterinary hospitals for diagnostic immunofluorescent antibody (IFA) and enzyme-linked immunosorbent assay (ELISA) testing of dogs suspected of a VBD. Dogs were suspected of a VBD on the basis of compatible historical and physical examination findings, as determined by the examining veterinary clinician. Available patient information included date of sample collection, date of sample receipt, animal signalment, and owner or veterinary practice address. Test results for this convenience sample were retrospectively reviewed and analyzed.
Serologic analysis
All serum samples included in the study were submitted to the VBDDL for evaluation using a defined canine VBD panel, which consisted of IFA testing against Brucella canis, Bartonella henselae, B. vinsonii subsp. berkhoffii, Ehrlichia canis, and R. rickettsii antigens. Additional testing for D. immitis antigen, antibodies to Anaplasma spp. (A. phagocytophilum and A. platys), Ehrlichia spp. (E. canis and E. chaffeensis), and the C6 peptide of B. burgdorferi were performed with a commercial ELISA-based kit (SNAP® 4DX®, IDEXX Laboratories, Westbrook, ME). To create a complete dataset, the SNAP® 4DX® test was performed retrospectively on samples submitted prior to 2007. Of note, no commercially available dog Lyme vaccine induces C6 peptide seroreactivity (O'Connor et al. 2004). With the SNAP® 4DX® assay, ELISA antibody cross-reactivity occurs with A. phagocytophilum and A. platys and also with E. canis and E. chaffeensis (Alleman et al. 2001, Otranto et al. 2009, Allison and Little 2013).
All IFA antigens were grown in vitro or, in the case of B. canis, in vivo by personnel in the VBDDL using bacterial or protozoan strains of canine or feline origin. Seroreactive samples were defined as having end point titers ≥1:64. B. henselae IFA was not performed in certain months of 2004, 2006, and 2007 due to a lack of antigen availability at the time of testing. By IFA testing, there is serological cross-reactivity between R. rickettsii antigens and antibodies directed against other SFG rickettsiae (including R. parkeri, R. felis, R. amblyommi, and other seemingly nonpathogenic SFG rickettsiae often found in ticks). There is also limited IFA cross-reactivity with E. chaffeensis and E. ewingii when testing against E. canis antigens. Therefore, IFA and ELISA seroreactivity, as reported in this study, indicates exposure to various Anaplasma, Ehrlichia, or SFG Rickettsia spp. Reactivity to E. canis by IFA or by ELISA was combined to represent exposure to one or more Ehrlichia spp. The SNAP® 4DX® and IFA tests do not distinguish between acute and chronic infections unless seroconversion is documented. Paired convalescent IFA titers documenting a four-fold rise in antibodies within a 4-week time frame support recent infection and a serological diagnosis of a specific VBD (Allison and Little 2013). Therefore, in this study, a sample submitted from the same dog within 5 weeks of initial testing likely represented a convalescent measurement and was defined as a duplicate sample. Duplicates were excluded from the database for analysis, as this study addressed exposure history, not documentation of active infection status.
Statistical analysis
Regions were based on address provided with sample submission and defined as follows: Northeast,CT, MA, ME, NH, RI, VT, NJ, NY, PA; Midatlantic, DE, DC, MD, VA, WV; South, NC, SC, GA, FL, KY, TN, AL, MS, OK, AR, LA, TX; Midwest, MI, OH, IN, IL, WI, MN, IA, MO, KS, NE, SD, ND; and West, MT, WY, CO, NM, ID, UT, AZ, NV, WA, OR, CA, AK, HI. Prevalence comparison analyses between regions and years were performed using the chi-squared or Fisher exact tests. Differences were considered significant at p≤0.05. Odds ratios (OR) and 95% confidence intervals (CI) by logistic regression were used to measure associations of co-exposures. In this study, the co-exposure OR (see Table 2, below) represents the odds of co-exposure to antigen 1 and antigen 2 as compared to the odds of exposure to a single antigen. Prevalence trends of each pathogen were evaluated using logistic regression modeling by day, month, and year of sample receipt. Graphic analysis included scatter plots with indicator coding, p test for linear trend, and seroreactivity point estimates with 95% confidence bands. Statistical analyses were performed using SAS/STAT 12.1 (SAS Institute Inc., Cary, NC).
Results
From 2004 through 2010, 14,496 US canine serum samples were submitted to the VBDDL for full VBD panel testing. Sixty-six duplicate submissions were identified and excluded from further analysis. Of the remaining 14,430 samples, the largest proportion originated from the South region (47.6%, n=6874), followed by the Midatlantic (20.4%, n=2945), Midwest (18.5%, n=2664), Northeast (7.7%, n=1108), and West (5.8%, n=839). Sample accessions received by state are presented in Table S1 (Supplementary Data are available at
Subspecies berkhoffi.
By immunofluorescent antibody.
By SNAP® 4DX®.
Anaplasma spp. prevalence, likely reflecting A. phagocytophilum exposure, was highest in the Northeast (7.5%; Table 1 and Fig. 1), with significant annual seroreactivity variation in the Midwest (p<0.01) and West (p=0.03). The annual seroreactivity of B. burgdorferi varied significantly over the 7-year study period in the Northeast (p=0.02), Midatlantic (p<0.01), Midwest (p<0.01), and South (p<0.01). Annual results of Borrelia seroreactivity were not significantly different between the Northeast and Midaltlantic regions over the duration of study (Fig. 2, p=0.5). Ehrlichia spp. seroreactivity was highest in the South and Midatlantic regions, with no significant difference in annual exposure trends between the two regions (p=0.9; Fig. 3).
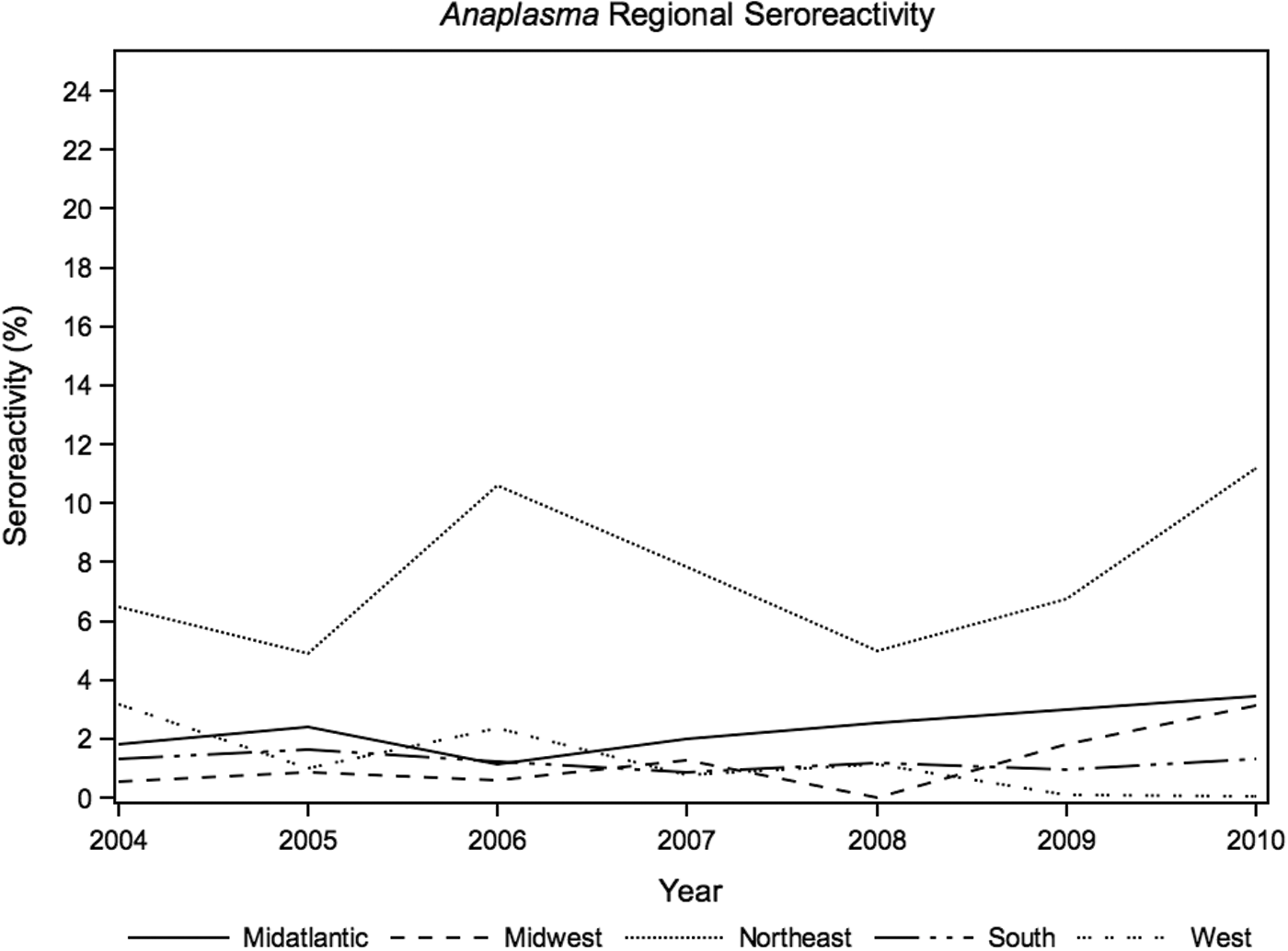
Annual canine seroreactivity of Anaplasma spp. by region, 2004–2010.
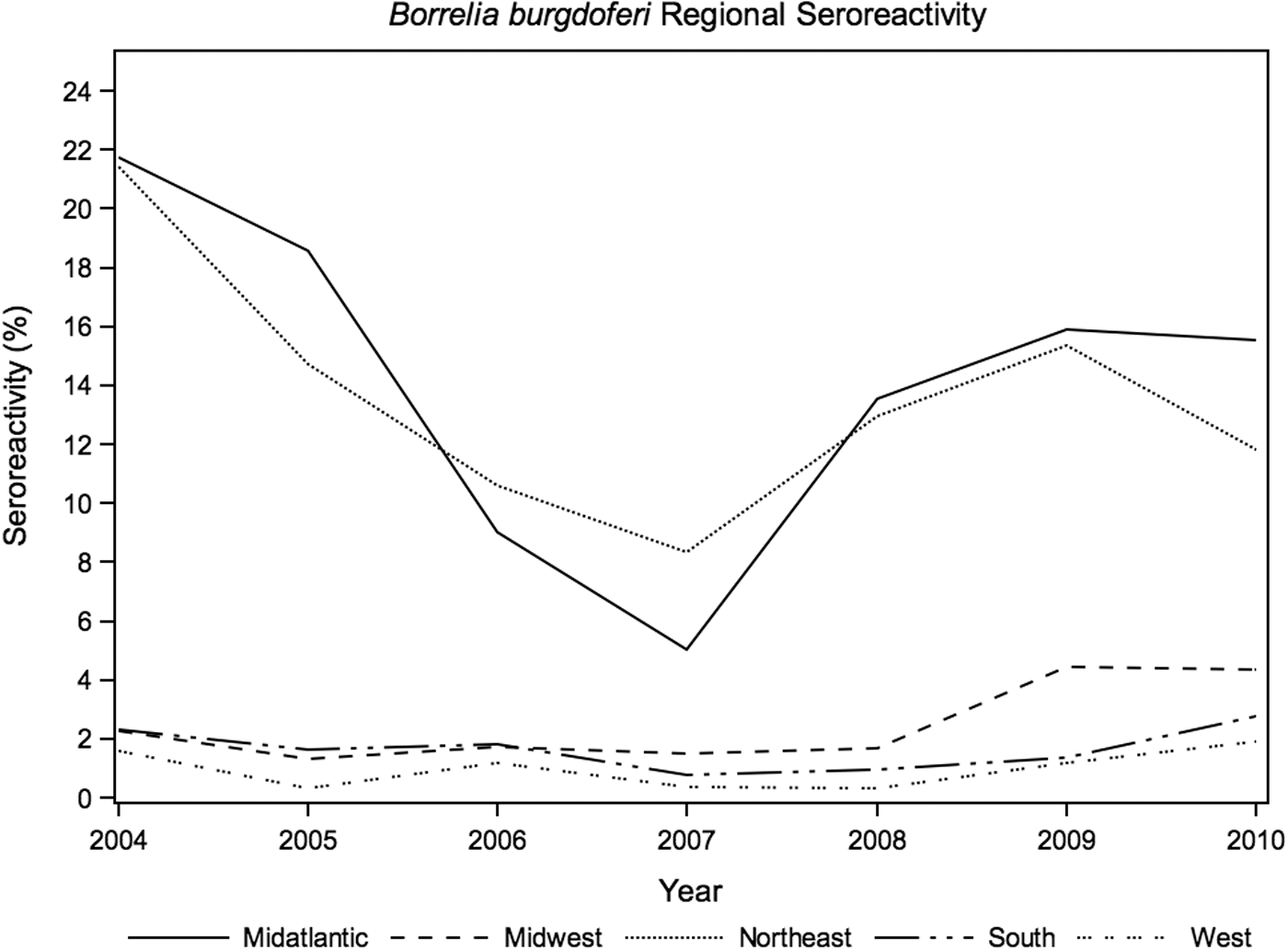
Annual canine seroreactivity of Borrelia burgdoferi by region, 2004–2010.
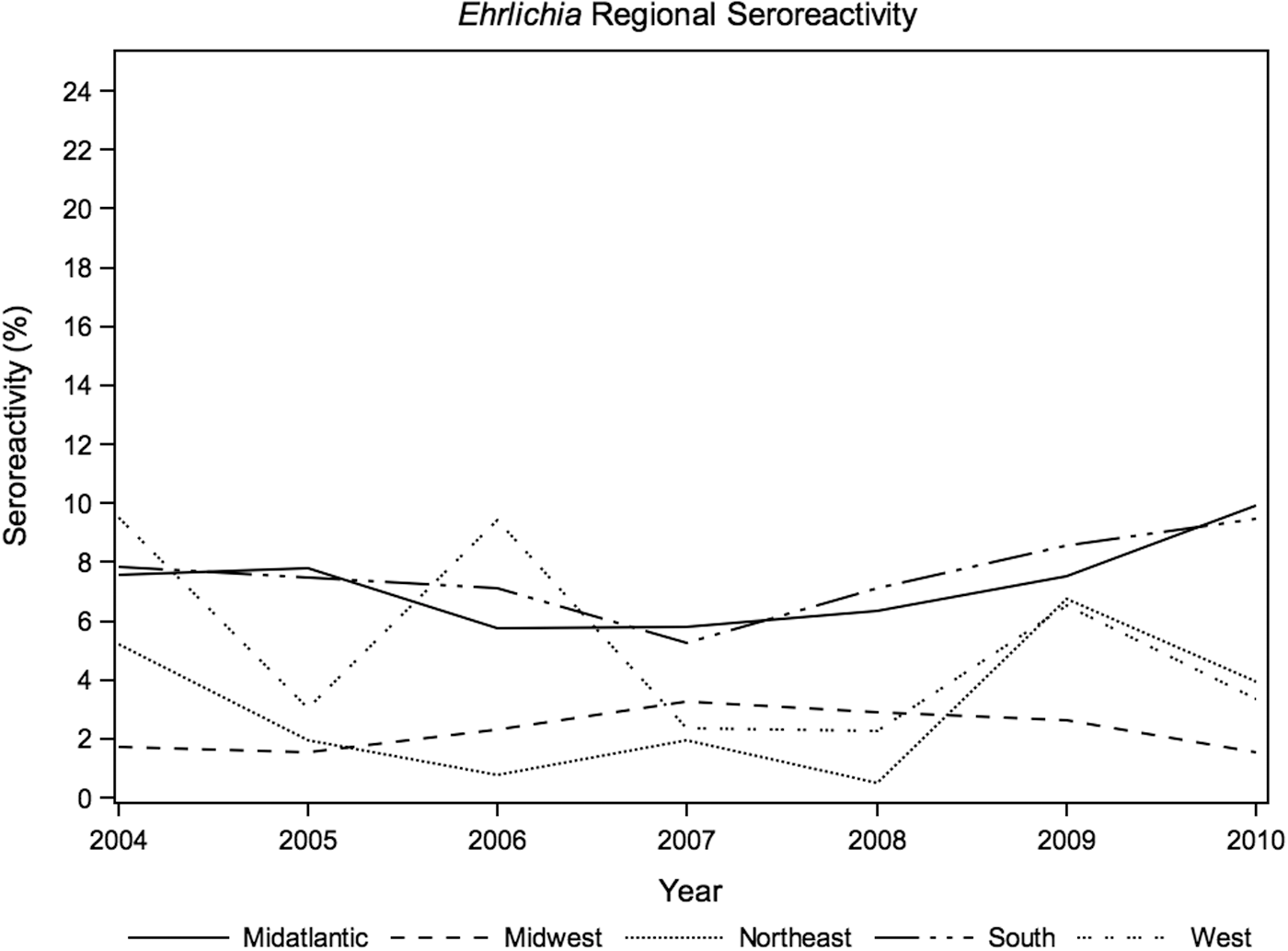
Annual canine seroreactivity of Ehrlichia spp. by region, 2004–2010.
SFG Rickettsia seroreactivity varied significantly by region (p<0.01), month (p<0.01), and year (p<0.01), with highest prevalence in the South and Midatlantic regions in 2005, and in the Midwest and Northeast in 2007 (Fig. 4). Rickettsia seroreactivity was higher in summer months, compared to other months within each year (Fig. 5). Annual seroprevalence changes of B. canis were significant in the Midwest (p≤0.01), South (p<0.01), and West (p=0.02), ranging from 0% to 3.5% over the study period (Fig. 6).
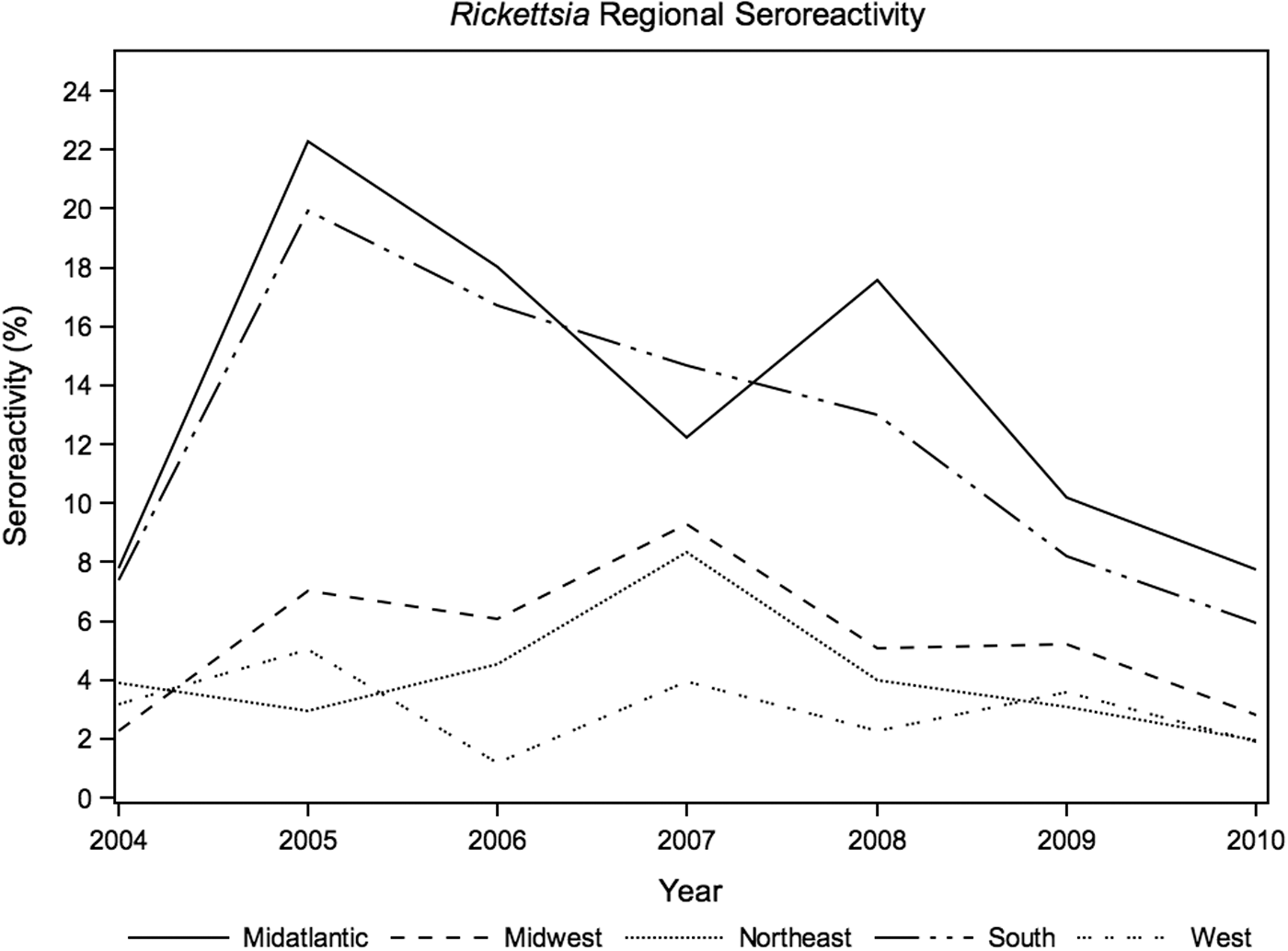
Annual canine seroreactivity of spotted fever group Rickettsia by region, 2004–2010.
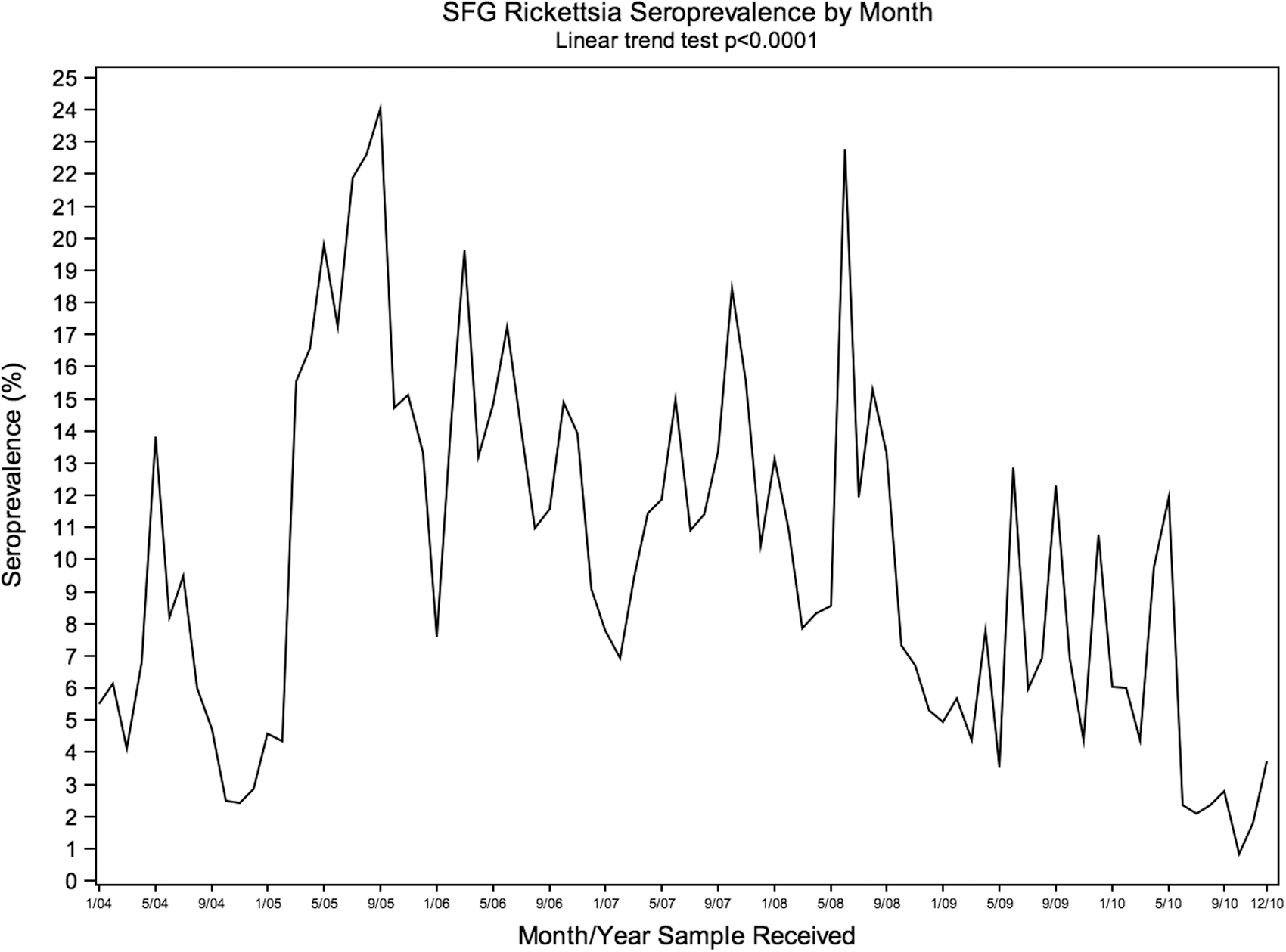
Monthly canine spotted fever group Rickettsia seroreactivity in the United States, 2004–2010. Month/year sample received.

Annual canine seroreactivity of Babesia canis by region, 2004–2010.
B. vinsonii subsp. berkhoffii exposure ranged from 0% to 3.4% over the duration of study (Fig. 7), with significant annual changes observed only in the South (p<0.01). D. immitis seroreactivity was less than 2% overall in the population and not observed in the West. Although B. henselae prevalence varied by year and region, missing data during periods of 2004, 2006, and 2007 decreases the validity of annual and regional comparisons and precluded monthly analysis of the data. No consistent monthly or seasonal variation in Anaplasma spp., B. burgdorferi, or Ehrlichia spp. seroreactivity was observed (data not shown). The annual seroreactivity of each pathogen by region is reported in Tables S2–S6.

Annual canine seroreactivity of Bartonella vinsonii subsp. berkhoffii by region, 2004–2010.
Of the 4278 seroreactive dogs in the United States, 704 (16.5%) were seroreactive to more than one antigen (Table 2). At the national level, combined seroreactivity to R. rickettsii in conjunction with Ehrlichia spp. was the most prevalent co-exposure (n=249, 5.8%), and the highest odds of co-exposure was observed with Anaplasma spp. and B. burgdorferi (OR=6.6, 95% CI 5.0, 8.8). Regionally, co-exposures were less prevalent in dogs from the Midwest and West, as compared to eastern US regions. Co-exposure antigens were regionally variable; the most prevalent antigens for each region are presented in Table 2.
Wald chi-squared test.
Discussion
Using a large diagnostic database, we analyzed the long-term exposure trends of eight VBD-causing pathogens in symptomatic dogs in the United States. Exposure to Anaplasma, spp., B. canis, B. henselae, B. vinsonii subsp. berkhoffii, B. burgdorferi, Ehrlichia spp., and SFG Rickettsia was identified in dogs from all US regions. Notable annual and regional seroreactivity variation of certain pathogens was observed over the years of study, consistent with findings in tick and human surveillance studies. Canine VBD seroprevalences have been correlated to human disease incidence, and with similar vector exposure as their owners, pet dogs can be effective sentinels for local VBD exposure risks for both animals and humans (Mead et al. 2011, Beall et al. 2012). With predictions of continued emergence of new tick-borne pathogens, consistent monitoring of dog and human populations are important components of VBD prevention and control (Nicholson et al. 2010). Our findings indicate canine serosurveys of selected VBD pathogens are capable of detecting prevalence changes over time, supporting the utility of canine surveillance systems in contributing to our knowledge of VBD risks.
In this population of dogs, for which a VBD was suspected, 29.6% were seroreactive to one or more of the evaluated antigens. We observed the highest overall exposure to Anaplasma in the Northeast, Borrelia in the Midatlantic and Northeast regions, and Rickettsia and Ehrlichia in the South, compatible with previous pet surveillance study findings (Bowman et al. 2009, Little et al. 2010, Beall et al. 2012). The seroreactivity to B. burgdorferi in NC and VA was higher in our study population than in an earlier 2001–2003 study (2.5% versus 0.4% in NC; 13.3% versus 8.7% in VA), using the same database and testing method (Duncan et al. 2005). The Borrelia prevalence in Maryland was stable between the two studies (14.9% and 14.4%) and had decreased in Pennsylvania (16.5% from 25.0% in 2001–2003). This may indicate a southern migration of the recognized vector of Lyme disease, Ixodes scapularis, from the Midatlantic states, or alternatively, a contribution of the southeastern Ixodes affinis vector in supporting B. burgdorferi transmission in the South (Maggi et al. 2010). Our findings for the Midatlantic region suggest the possibility of pathogen convergence, with the highest overall regional seroreactivity (43.4%), and a comparatively high prevalence of pathogens historically considered regionally localized, including Northeast-endemic pathogens A. phagocytophilum and B. burgdorferi, and South-endemic pathogens Ehrlichia and Rickettsia spp.
The seroreactivity to B. burgdorferi, Ehrlichia spp., and SFG Rickettsia demonstrated significant annual and regional variation among this study population. Annual seroreactivity trends for each pathogen were region specific. Although Anaplasma spp. and B. burgdorferi share a similar vector–reservoir–host system (Nicholson et al. 2010), the annual variation in trends did not coincide between the two antigens in the Northeast. SFG Rickettsia demonstrated consistent seasonal seroreactivity variations, as has been observed with human surveillance of RMSF cases (Openshaw et al. 2010). Significant seasonal variation was not observed for the remaining study pathogens. Because canine VBDs can have incubation periods ranging from weeks to months, attempting to correlate disease diagnosis data with pathogen exposure and vector-related seasonal trends may be difficult or implausible. Additional study is warranted to explore factors that contribute to observed VBD seroprevalence variations in this and other dog populations.
Among the exposed dogs in this study, co-exposure prevalence of the eight pathogens was moderate (16%), with high odds of co-exposures for some pathogens and in some regions. Several studies are available on tick-borne co-infections in tick, human, and wildlife populations, but co-exposure prevalence data in domestic animals in the United States is limited (Nieto and Foley 2009). In our study, the highest odds of a co-exposure were observed with pathogens that share similar vectors; odds were lowest for co-exposures with pathogens that generally have different vectors, including Rickettsia with Borrelia and Ehrlichia with Borrelia. The pathogenesis resulting in various co-infections is not well established; pathogen synergism has been proposed and alterations of the host immune response have been found experimentally (Nieto and Foley 2009, Gaunt et al. 2010). Long-term antibody persistence of select VBD pathogens, including Ehrlichia and Rickettsia, contributes to the challenges of co-exposure interpretation. In light of the detrimental clinical impact associated with VBD co-infections, further characterization of co-exposure epidemiology would benefit both animals and humans.
Several factors should be considered in the assessment of our findings. New acaracide products, with improved tick efficacy, were introduced onto the market over the years of this study, resulting in increased marketing and availability of these products by dog owners. Additionally, veterinarians had increasing accessibility to VBD educational resources and novel testing modalities. Collectively, these factors contributed to increased awareness of VBDs among veterinarians, resulting in increased VBD testing of sick dogs, and increased accessibility to tick preventative and Lyme disease vaccine strategies. In addition, national and international pet travel and animal relocation are more commonplace now than at any time in history, which, along with intentional and unintentional wildlife movements, may introduce VBDs into new geographic regions and contribute to the challenges in accurately defining the local and regional epidemiology of VBDs in dogs.
Several limitations may have influenced our findings. Due to clinical suspicion of a VBD, this study population originated from clinician-submitted samples, creating a potential bias for higher VBD seroprevalence. This sampling limits the generalizability of the study findings to the healthy US dog population or to dogs with limited veterinary care. Additional factors that may reduce accuracy or bias our results include: The South region was overrepresented in the study population; the SNAP® 4DX® test is intended as a screening test, therefore does not consistently detect low E. canis antibody titers (IFA titers <1:256); some diagnostic samples were submitted from regional referral hospitals, therefore zip code data may not accurately reflect the dogs' original exposure environment; and lack of a travel history compromises confirming the site(s) of vector exposure. On the basis of comparison studies using PCR, Bartonella serology results may have grossly underestimated exposure in the dogs of this study (Perez et al. 2011).
Conclusions
We report the results of the first long-term survey of exposure to Anaplasma spp., B. canis, Bartonella spp., B. burgdorferi, D. immitis, Ehrlichia, and Rickettsia spp. in dogs from the United States. Notable annual and regional seroprevalence variation was observed with certain pathogens over the 7 years of study, suggesting canine surveillance studies may have value in contributing to our future VBD knowledge. A multidisciplinary approach to further characterize the epidemiology of VBD pathogens can lead to more effective prevention strategies and improved health outcomes for dogs and humans.
Footnotes
Acknowledgments
We thank the personnel of the Vector Borne Disease Diagnostic Laboratory, Monica Morgado and Natalie Cherry for performing the testing and IDEXX Laboratories Inc, Westbrook, ME, for providing the SNAP® 4DX® assays used in this study. This research was supported in part by the State of North Carolina.
Author Disclosure Statement
Adam J. Birkenheuer, Barbara C. Hegarty, Michael G. Levy, Pedro P.V.P. Diniz, David J. Weber, and Caroline B. Yancey have no competing financial interests. Edward B. Breitschwerdt is a consultant for IDEXX Laboratories and is the Chief Medical Officer for Galaxy Diagnostics. Barbara A. Qurollo is a postdoctoral research fellow in the Intracellular Pathogens Research Laboratory and funded by IDEXX Laboratories.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.