
Abstract
Epidemiological studies worldwide have reported a high prevalence and a great diversity of Bartonella species, both in rodents and their flea parasites. The interaction among Bartonella, wild rodents, and fleas reflects a high degree of adaptation among these organisms. Vertical and horizontal efficient Bartonella transmission pathways within flea communities and from fleas to rodents have been documented in competence studies, suggesting that fleas are key players in the transmission of Bartonella to rodents. Exploration of the ecological traits of rodents and their fleas may shed light on the mechanisms used by bartonellae to become established in these organisms. The present review explores the interrelations within the Bartonella–rodent–flea system. The role of the latter two components is emphasized.
Introduction
T
Rodents have been found to be natural reservoirs of many Bartonella species. The association between rodents and bartonellae is of great importance, because the former display persistent and subclinical bacteremia that lasts for months. In addition, many different rodent species have been reported to be infected in high percentages with different Bartonella spp. and variants worldwide (Fig. 1).
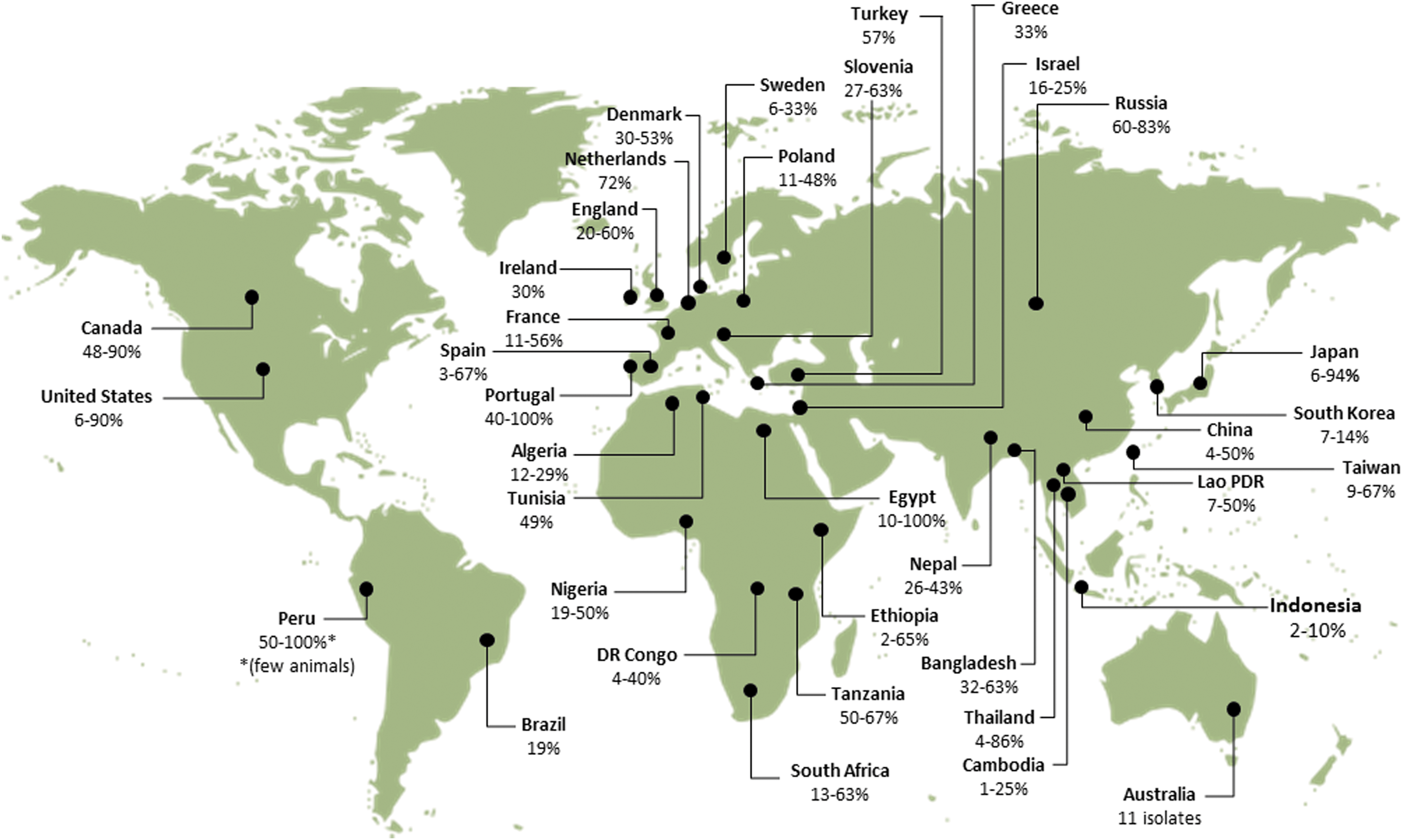
Range of Bartonella-infection rates in rodents reported worldwide (data from PubMed, January, 2014). References according to continents and countries: Africa, Algeria, Bitam et al. 2009; Egypt, Inoue et al. 2009; Ethiopia, Meheretu et al. 2013; DR Congo and Tanzania, Gundi et al. 2012b; Nigeria, Kamani et al. 2013; South Africa, Pretorius et al. 2004, Brettschneider et al. 2012b, Trataris et al. 2012; Tunisia, Fichet-Calvet et al. 2000; Asia, Bangladesh, Bai et al. 2007b; Cambodia, Lao PRD, and Thailand, Jiyipong et al. 2012; China, Ying et al. 2002, Ye et al. 2009, Inoue et al. 2009, Liu et al. 2010; Indonesia, Winoto et al. 2005; Israel, Harrus et al. 2009, Morick et al. 2009; Japan, Inoue et al. 2008, Kabeya et al. 2011; Lao PDR, Angelakis et al. 2009; Nepal, Gundi et al. 2010; Taiwan, Lin et al. 2008, Hsieh et al. 2010, Tsai et al. 2010, Chae et al. 2008; Russia (Far east), Mediannikov et al. 2005; South Korea, Kim et al. 2005; Thailand, Castle et al. 2004, Bai et al. 2009, Saisongkorh et al. 2009, Inoue et al. 2009; Turkey (Kaman, Kirsehir), Karagöz et al. 2013; America, Brazil, Costa et al. 2014; Canada, Jardine et al. 2005, Jardine et al. 2006b; Peru, Birtles et al. 1999; United States, Kosoy et al. 1997, Ellis et al. 1999, Bown et al. 2002, Kosoy et al. 2003, 2004a, Bai, et al. 2007a, 2008b, Morway et al. 2008, Matsumoto et al. 2010, Bai et al. 2011, Gundi, et al. 2012a; Europe, Denmark, Engbaek and Lawson 2004; England, Birtles et al. 2001, Bown et al. 2002, Telfer et al. 2007a, France, Gundi et al. 2004, Buffet et al. 2012, 2013b; Greece, Tea et al. 2004; Ireland, Harrison et al. 2012; Netherlands, Holmberg et al. 2003; Poland, Welc-Faleciak et al. 2008, 2010, Paziewska et al. 2012a, Hildebrand et al. 2013; Portugal, Ellis et al. 1999; Slovenia, Knap et al. 2007; Spain, Marquez et al. 2008, Gil et al. 2010; Oceania, Australia, Gundi et al. 2009. Color images available online at
Fleas are considered key players in the Bartonella cycle, because they harbor a high diversity of Bartonella spp. and strains and demonstrate great efficiency in the transmission of these bacteria among rodents (Brinkerhoff et al. 2010, Morick et al. 2013a). Therefore, fleas were suggested not to serve solely as vectors, but to represent additional reservoirs for these bacteria (Birtles 2005, Deng et al. 2012).
Several rodent-associated Bartonella spp. and variants have been implicated as the causative agents of human clinical manifestations, including endocarditis (Daly et al. 1993, Fenollar et al. 2005), myocarditis (Kosoy et al. 2003), fever and neurologic disorders (Welch et al. 1999), intraocular neuroretinitis (Kerkhoff et al. 1999), meningitis (Probert et al. 2009), splenomegaly (Eremeeva et al. 2007), and lymphadenopathy (Oksi et al. 2013). These findings make rodents and their fleas a relevant natural system for the study of ecological pathways of vector-borne pathogens and emerging diseases of human importance.
This review attempts to summarize and bridge some knowledge gaps in the transmission and distribution routes and in the dynamics and composition of Bartonella infection in rodents and their flea parasites. Many studies and reviews have focused on the bacterial component within the organism–host–vector triangle (Saenz et al. 2007, Chomel et al. 2009, Vayssier-Taussat et al. 2010, Engel et al. 2011, Deng et al. 2012, Harms and Dehio 2012, Buffet et al. 2013a, Buffet et al. 2013b). In this review, we focus on the particular ecological traits of the other two components, the rodent and the flea, that may explain the apparent co-adaptation of these organisms.
Bartonella in Rodents and Their Associated Fleas
Rodents infected with Bartonella spp. have been described since the mid-twentieth century (Baker 1946, Kosoy 2010), but the majority of reports on Bartonella-infected rodents have significantly accumulated after the expansion of the Bartonella genus in 1993 (Kosoy 2010). Bartonella spp. have been virtually detected from rodents worldwide (Fig. 1). Pioneering studies were carried out in North America and Europe, and later in Asia, Africa, Latin America, and Oceania (Fig. 1). Extremely high Bartonella infection rates were reported in rodent communities reaching up to 90.4% in the northern grasshopper mice (Onychomys leucogaster) from Kansas (Bai et al. 2007a) and 82.4% in the deer mice (Peromyscus maniculatus) from Colorado (Bai et al. 2011), suggesting a mutual adaptation between the bacteria and their reservoirs. Yet, some species have shown to be less susceptible or completely resistant for infection. For instance, only seven of 14 rodent species captured in grasslands of four different states in the United States were found to be infected (Bai et al. 2007a).
Rodents harbor the greatest diversity of bartonellae described to date. Numerous isolations of Bartonella spp. and variants have been obtained from more than 98 rodent species belonging to at least seven families. About 22 rodent-associated Bartonella spp. and subspecies have been described, some of which were initially isolated from human cases (Buffet et al. 2013a). The chronological succession of the description of these Bartonella spp. is: B. elizabethae and B. vinsonii subsp. vinsonii (Brenner et al. 1993), B. doshiae, B. grahamii, and B. taylorii (Birtles et al. 1995), B. tribocorum (Heller et al. 1998), B. vinsonii subsp. arupensis (Welch et al. 1999), B. birtlesii (Bermond et al. 2000), B. washoensis (Kosoy et al. 2003), B. phoceensis and B. rattimassiliensis (Gundi et al. 2004), B. rochalimae (Lin et al. 2008), B. tamiae (Kosoy et al. 2008), B. rattaustraliani, B. queenslandensis, and B. coopersplainsensis (Gundi et al. 2009), B. japonica and B. silvatica (Inoue et al. 2010), and B. jaculi, B. callosciuri, B. pachyuromydis, and B. acomydis (Sato et al. 2013). Other Bartonella isolated from rodents have been proposed as new species or subspecies, including Candidatus Bartonella washoensis subsp. cynomysii (Bai et al. 2008a), Candidatus Bartonella volans, Candidatus Bartonella durdenii, and Candidatus Bartonella monaxi (Breitschwerdt et al. 2009). Phylogenetic analysis of the characterized species has shown that the majority of Bartonella spp. are clustered in a common lineage, with the exception of B. rochalimae and B. tamiae (Buffet et al. 2013a), evidencing the occurrence of an adaptive evolution of these bacteria in rodents (Engel et al. 2011). Moreover, several molecular studies have found a wider variety of Bartonella genotypes that have challenged the current taxonomic classification (Bai et al. 2009, Harrus et al. 2009, Inoue et al. 2009). To clarify the vast rodent–bartonellae diversity, the use of alternative taxonomic classification according to species complexes has been proposed (Kosoy et al. 2012). The recent elevated number of reported new Bartonella spp. and genotypes from rodents can be explained by a potential accelerated evolution of the Bartonella genus in rodents as a result of frequent recombination events, horizontal gene acquisitions, and accumulation of mutations (Berglund et al. 2010, Guy et al. 2013). Increased Bartonella-oriented research can also contribute to this phenomenon.
Bartonella infection in rodents can be composed of more than one Bartonella sp. or genetic variants in the same rodent (i.e., co-infection) (Morick et al. 2011). The potential interaction between the co-existing Bartonella variants could be the source of the recombination events and the diversity revealed in rodent-associated bartonellae (Berglund et al. 2009, Berglund et al. 2010, Paziewska et al. 2011, Paziewska et al. 2012b). Different rodent or flea species can play a greater role in the occurrence of such events (Paziewska et al. 2012a). In addition, evidence for the presence of the same Bartonella sp. in two different rodent species has been accumulated and described as the “spillover” phenomenon (Ying et al. 2002, Castle et al. 2004, Kosoy et al. 2004b, Jardine et al. 2006a, Bai et al. 2007a, Telfer et al. 2007b). This phenomenon can be driven by the act of ectoparasites exchanged between different rodent species or even genera, or by close interspecies and/or intergeneric interaction between rodents.
Among the ectoparasites infesting rodents, fleas are considered major vectors of bartonellae (Billeter et al. 2008, Tsai et al. 2011). Various fleas have been demonstrated to acquire and transmit Bartonella spp. and strains under experimental conditions, probably serving as competent vectors of these bacteria in the wild (Krampitz 1962, Bown et al. 2004, Morick et al. 2011, Morick et al. 2013a). Bartonellae have been shown to be dominant members of the bacterial communities of several rodent associated fleas (Jones et al. 2008, Hawlena et al. 2013). Moreover, many rodent-associated fleas have been shown to be naturally infected with Bartonella closely related to zoonotic species (Stevenson et al. 2003, Marie et al. 2006, Jones et al. 2008, Morick et al. 2010, Billeter et al. 2011, Kabeya et al. 2011, Billeter et al. 2013, Kim et al. 2013). The frequent feeding of fleas and their ability to move from one host to another might explain the high prevalence and diversity of Bartonella spp. infection found in rodents and fleas (Kosoy et al. 2012). Moreover, adaptation between bartonellae and fleas was evidenced through the experimental infection of Xenopsylla ramesis fleas with Bartonella sp. OE 1-1, a strain closely related to B. elizabethae. Infection with this Bartonella variant did not affect the metabolic rate, blood consumption, life span, fertility, or fecundity of female fleas, nor the developmental time, the life span, or sex ratio of their offspring fleas (Morick et al. 2013b). Interestingly, bartonellae DNA has been detected in other rodent ectoparasites, including ticks, mites, and lice (Durden et al. 2004, Kim et al. 2005, Reeves et al. 2006). A recent study has demonstrated the competency of Ixodes ricinus ticks to transmit the rodent-associated B. birtlesii (Reis et al. 2011). However, the biological role of ticks in transmission of Bartonella in nature is still under debate, and their epidemiological role has been considered secondary (Matsumoto et al. 2010, Harrison et al. 2012). In this review, the role of fleas in the ecology of bartonellae in rodents is emphasized, although alternative vectors may also play a role in the transmission cycle.
Ecological Insights of the Rodent-Flea-Bartonella Triangle
Better understanding of rodent ecological traits and the dynamics of flea infestation may shed light on the pathways that enabled bartonellae to be established in rodent hosts and flea vectors. Rodents' habitat, behavior, and flea parasitism may represent crucial variables influencing the transmission and establishment of bartonellae, and these typically vary across species and geographical areas.
The rodent habitat can represent an important variable in the efficiency of the transmission cycle and in the selection of Bartonella spp. repertoire to which rodents are exposed. The burrow habitat can directly affect the flea cycle, because immature stages live off-host and exhibit certain microclimatic preferences (Krasnov et al. 2001). Therefore, if the conditions are not favorable for flea development, vectorial transmission of bartonellae may not occur. The habitat location and geographic conditions (e.g., rainy versus dry seasons) can also influence the feeding performance of certain rodents and can lead to an increased vulnerability for bacterial infections (Beldomenico and Begon 2010), especially for those who base their nutrition on green vegetation. Visit of burrows or invasion to the territories of other rodent species can represent an important source of flea interchange (Krasnov and Khokhlova 2001). Therefore, the geographical and/or habitat traits of rodents and their fleas may enhance or restrict the diversity of bartonellae observed in those animal communities (Jardine et al. 2006a, Morick et al. 2010).
Rodent behavior can directly influence the transmission of bartonellae between members of the same community. Physical contact between individual rodents may promote the transmission of bartonellae. Grooming can facilitate the acquisition of bartonellae by disrupting the skin barrier (e.g., by aggressive grooming) or by removal and interchange of ectoparasites between rodents (e.g., social grooming) (Krasnov and Khokhlova 2001, Stopka and Graciasova 2001). Parental behavior (carrying, licking, and huddling of young animals) (Lonstein and De Vries 2000) can lead to an early exposure and transmission of bartonellae among rodents. In addition, mobility, spatial behavior, and seasonality of the rodents (e.g., for reproductive purposes or emergence of juveniles from burrows) are traits that can contribute to the risk of bartonellae acquisition in wild rodents by influencing the rate of rodent–rodent interactions (Krasnov et al. 2005). In fact, during reproductive periods, male rodents tend to increase their mobility and may suffer immunosuppression due to effects of sex hormones (Krasnov et al. 2005) that can lead to a major susceptibility to bacterial infections. Moreover, hibernation can also play a role in the persistence or clearance of infection in rodent communities across seasons (Jardine et al. 2006b). Nevertheless, all of these particular behaviors can vary across rodent species and may reflect the different host adaptability to bartonellae of certain rodents.
Flea parasitism likely plays a fundamental role in the transmission and acquisition of bartonellae being mediated by the level of flea's host specificity, flea exchange between rodents, and flea abundance. The host specificity of fleas is an important trait that can influence the introduction or the restriction of new Bartonella spp. and strains to new hosts. Fleas vary greatly in the degree of their host specificity, from being highly host specific to host opportunistic (Krasnov et al. 2004). The latter will underline the potential success of exchanging fleas between rodents. In fact, the exchange of fleas can likely occur between conspecific rodents or even across rodent species in the wild (Krasnov and Khokhlova 2001). The flea abundance (number of fleas per host) might directly influence the chances of bartonellae infection establishment in the host. It is noteworthy that the abundance of fleas varies across rodent species, flea species habitats, and seasons (Krasnov et al. 1997, Krasnov et al. 2005, Kim et al. 2013).
Acquisition and Transmission Pathways of Bartonella
Investigation of the transmission routes and dispersal dynamics of bartonellae among animals and their vectors is crucial in elucidating the ways they are being maintained in nature. Microorganisms are transmitted via vertical and/or horizontal pathways to spread within host populations. Thereby, competence studies have contributed greatly to the comprehension of rodent-associated bartonellae cycles (Fig. 2).
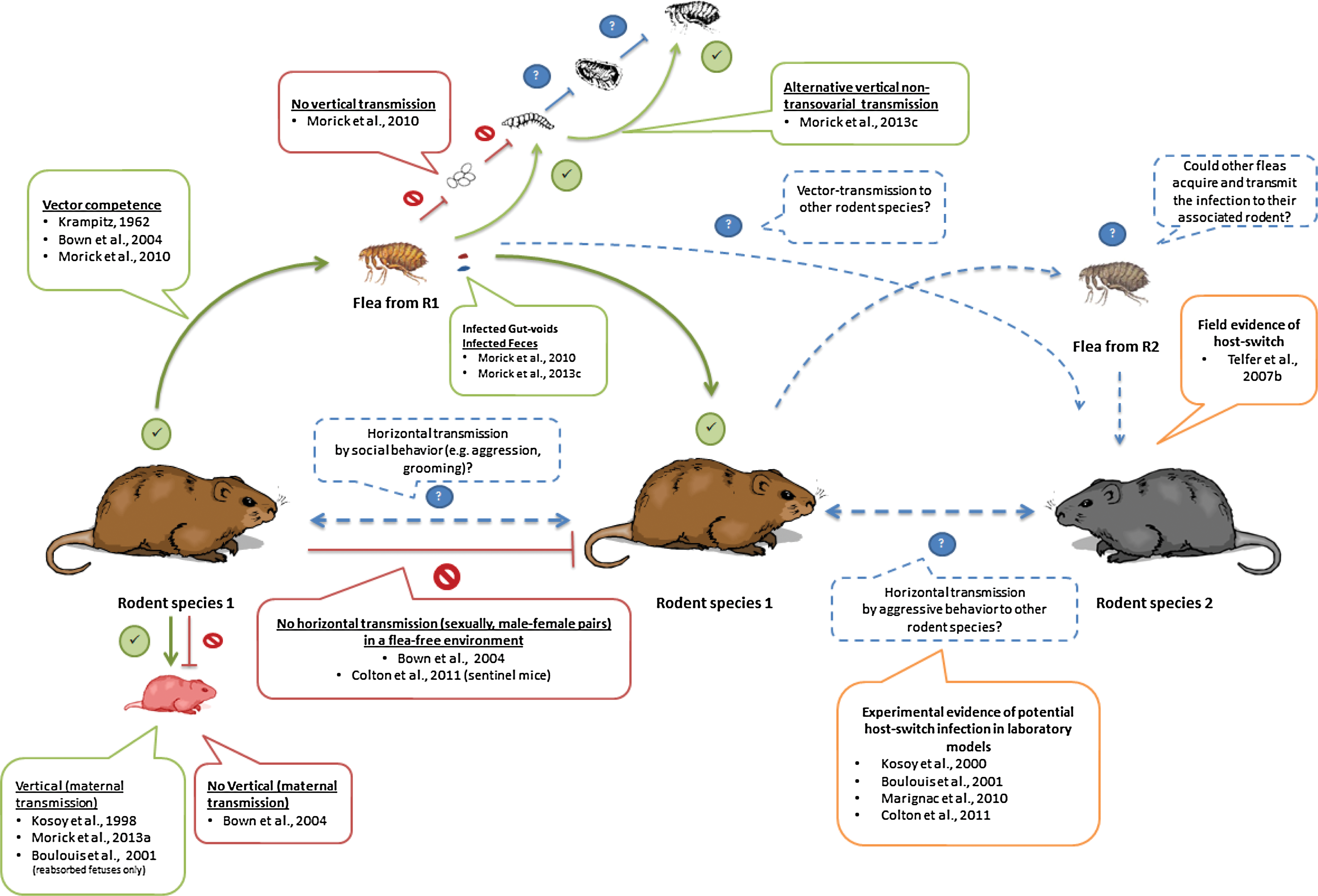
Experimentally demonstrated pathways and potential transmission routes of bartonellae in wild rodents and their fleas. (Green lines) Experimentally demonstrated pathways; (red lines) investigated routes that failed to be demonstrated; (dashed blue lines) proposed transmission pathways. Boxes include the corresponding reference studies. Yellow-lined boxes indicate indirect evidence for the suggested pathways. Color images available online at
Various studies have demonstrated the capability of fleas to acquire and transmit Bartonella strains under experimental conditions. Xenopsylla cheopis fleas were reported as competent vectors of an unidentified Bartonella sp. on Myodes glareolus voles through a pioneer study (Krampitz 1962). More recently, wild-collected Ctenophthalmus nobilis demonstrated the ability to transmit B. grahamii and B. taylorii to captive-bred M. glareolus voles (Bown et al. 2004) and Xenopsylla ramesis has been shown to be a competent vector of Bartonellae sp. OE 1-1 to Meriones crassus jirds (Morick et al. 2011, Morick et al. 2013a). The latter studies demonstrated a remarkable efficiency of fleas in acquiring and transmitting Bartonella. About 69%–100% of the fleas acquired the Bartonella sp. within a period of 72 h, and a similar period was sufficient for the fleas to infect naïve jirds. It is acknowledged that this transmission route from arthropod to mammal is done through the gastrointestinal content, mainly feces (stercoraria) (Birtles 2005). Supporting evidence for this route was initially demonstrated in cats infected through the inoculation of cat-flea (Ctenocephalides felis) feces (Foil et al. 1998). As stercoraria may seem an inefficient infection route (Birtles 2005, Marignac et al. 2010), further studies are required to investigate the occurrence of stercoraria, determine possible fecal components that may promote entry and establishment of bartonellae in the intradermal compartment, calculate the amount of fecal depositions in host skin per a specific time period, and ultimately define the minimal bacterial load required for infection efficiency.
Alternative vertical or horizontal transmission pathways of Bartonella infection among rodents, without the mediation of an arthropod, have been studied under natural and laboratory conditions. First, vertical transmission in wild-captured Sigmodon hispidus and Peromyscus leucopus rodents was observed by isolating Bartonella spp. from the placental tissues of the pregnant animals, embryos, and from neonatal pups (Kosoy et al. 1998). Later, Boulouis et al. (2001) explored the vertical transmission of B. birtlesii in BALB/c mice. The authors obtained similar results in the transplacental transmission of B. birtlesii in bacteremic pregnant BALB/c mice. However, isolation of the bacteria was possible only from reabsorbed fetuses, but not from dead or viable ones. Contrary to those studies, no transmission was detected from infected females to their offspring in bank voles (M. glareolus) (Bown et al. 2004). Recent studies on a desert rodent, M. crassus, demonstrated the presence of Bartonella DNA in one of 15 pups born to experimental infected jirds (Morick et al. 2013a). Altogether, these studies suggest that this maternal transmission route is feasible, although it seems to have a minor role in the Bartonella cycle in the wild. Horizontal transmission between male and female rodents was also investigated, where rodents were kept under an arthropod-free environment, resulting in no transmission of Bartonella from the experimentally infected to the naïve rodents (Bown et al. 2004). To fill the knowledge gaps in arthropod-free transmission routes, studies evaluating the potential interaction (e.g., social or aggressive interactions) between rodents from the same sex and between different species and genera are required (Fig. 2).
Arthropods may serve as reservoirs for bacteria in nature. To accomplish such role, the arthropod needs to allow the transmission of the bacteria from one generation to another through vertical mechanisms, such as transstadial, transovarial, co-feeding, or even sexual mechanisms (Parola and Raoult 2001). The vertical transmission of Bartonella among arthropods has been explored in various studies (Halos et al. 2004, Cotte et al. 2008, Morick et al. 2011, Morick et al. 2013c). The detection of Bartonella DNA in the reproductive tissues of Cediopsylla inaequalis, Oropsylla hirsuta, Aetheca wagneri, and Orchopeas leucopus (Brinkerhoff et al. 2010), and Ctenophthalmus congener truncus and Neopsylla sasai (Kabeya et al. 2011) fleas, highlighted the potential transovarial transmission of Bartonella in fleas. However, X. ramesis–infected fleas did not show a transovarial transmission to their offspring (Morick et al. 2011). It has to be acknowledged that flea interspecies differences might be involved in such contrasting results; thus, this route cannot be ruled out for all flea species. Additionally, in the latter study, the presence of Bartonella DNA in gut voids from infected fleas suggested an alternative non-transovarial transmission route, since it is known that underdigested blood regurgitated by parent fleas can serve as food supply for flea larvae (Krasnov 2008, Khokhlova et al. 2010). This route was later demonstrated through the acquisition of Bartonella infection by immature flea stages that were exposed to gut voids and/or feces deposited by infected adult fleas (Morick et al. 2013c). Similarly, other arthropods have shown this non-transovarial transmission route: The stinkbug Riptortus clavatus were shown to acquire its Burkholderia sp. gut symbiont from the environment on nymph stages, allowing the transfer of the symbiont every generation through a postnatal mechanism (Kikuchi et al. 2007). This transmission route gives extra significance to the microhabitat in burrows of rodents for the continuous infection of bartonellae among fleas. It can be hypothetized that old and abandonned rodent burrows can still serve as potential temporal reservoirs of arthropods for bartonellae infection through the presence of flea feces or gut voids that can serve as food for newly hatched flea larvae. Nevertheless, further questions, such as the time length that these gut contents remain infective, have to be answered to verify this hypothesis.
Alternative routes of infection have been explored in mammals. Guptill et al. (1999) reported infection through oral inoculation of B. henselae in cats. Foil et al. (1998) explored additional infection strategies of B. henselae in cats, including bacteremic blood injections. Similarly, alternative routes were tested in rodents, including oral and ocular routes (Marignac et al. 2010). These experiments investigated the potential acquisition of Bartonella infection by ingestion of fleas by the rodents (e.g., as a result of grooming) or the direct inoculation of the bacteria into the eyes (e.g., as a result of aggression behavior) that could prompt these membranes as alternative entry pathways. Nevertheless, these routes showed to be less efficient and required higher doses to produce bacteremia.
Bartonella Infection Dynamics: From Experimental Infection to Field Studies
Experimental infection
The infection dynamics of rodent-associated bartonellae have been extensively explored and partially elucidated in wild-captured and laboratory rodent models (Kosoy et al. 1999, Kosoy et al. 2000, Boulouis et al. 2001, Koesling et al. 2001, Schulein et al. 2001, Marignac et al. 2010, Colton et al. 2011, Colton and Kosoy 2012, Morick et al. 2013a). The first phase of the Bartonella infection, spanning from the inoculation of the flea infected-contents (e.g., feces and gut voids) on the wounded skin of the mammal to the appearance of the bacteremia still remains obscure (Chomel et al. 2009, Harms and Dehio 2012). An incubation period has been evidenced after rodents were inoculated with bartonellae by intravenous, subcutaneous, intradermal, or flea-infected challenges (Schulein et al. 2001, Marignac et al. 2010, Morick et al. 2013a). Thus, a primary niche for the infection has been proposed (Chomel et al. 2009, Harms and Dehio 2012). Although, nonconclusive in vivo evidence of this niche or the potential cells involved have been published to date, the vascular endothelial cells seem to be the major candidate for this niche (Deng et al. 2012). Once the bartonellae reach the bloodstream, they infect their final target cells, the erythrocytes. Schulein et al. (2001) observed cyclic bacteremic waves during the course of infection after intravenous challenge, supporting the role of the proposed niche in reseeding the bacteria into the bloodstream.
In a recent study, bacteremic waves were observed in naturally infected jirds, and in jirds challenged with infected fleas or subcutaneous inoculation, raising supporting evidence that this phenomenon occurs in the wild (Morick et al. 2013a). It has been proposed that these cyclic bacteremic waves are the result of an apparent clearance of the bacteria by the immune system, followed by a new bacterial input from the primary niche to the bloodstream (Harms and Dehio 2012). However, because infection with Bartonella does not cause hemolysis or reduced erythrocytes life span, it is likely that those reinfection waves are a product of lytic cycles of the primary niche (Schulein et al. 2001). Interestingly, bacteremia relapses were not reproducible in outbred and inbred heterologous mice models, even after induction of immunosuppression (Marignac et al. 2010). Therefore, if the primary niche is playing the proposed role, this may occur only in highly adapted hosts (natural reservoir). Through this unique infection strategy, bartonellae have evolved in a way that permits hematophagous arthropods to efficiently acquire the bacteria and successfully transmit it to other hosts (Schulein et al. 2001, Chomel et al. 2009).
Infection dynamic studies using murine models infected with adapted bartonellae have reproduced long-lasting bacteremia under laboratory conditions. Cotton rats (S. hispidus) infected with three Bartonella strains isolated from the same rodent species, reproduced bacteremia for up to 15 weeks (Kosoy et al. 1999). Notably, two Sundevall's jirds (M. crassus) naturally infected with Bartonella sp. OE 1-1, kept under laboratory conditions, remained bacteremic for 24 and 30 weeks, respectively. Additionally, subcutaneously and flea-challenged naïve rodents showed infection persistence of up to 23 weeks (Morick et al. 2013a). These results in rodent models mirrored the patterns observed in other natural reservoirs, such as cats infected with B. henselae (Kordick et al. 1995).
To evaluate the course of Bartonella infection in accidental hosts, murine laboratory animals were challenged with Bartonella strains isolated from other rodent species. Interestingly, these studies have raised contradicting results. Through an extensive challenge study, using cotton rats (S. hispidus), white-footed mice (P. leucopus), BALB/c (Mus musculus), and Wistar rats (Rattus norvegicus) inoculated with 14 Bartonella strains (including human isolates), Kosoy et al. (2000) observed that bacteremia could be reproduced only in cotton rats and white-footed mice infected with Bartonella strains isolated from the same species or from congenic rodents. In contrast, other studies have reproduced long-lasting bacteremia (up to 11 weeks postinfection [p.i.]) in laboratory mice (M. musculus inbred strains) with B. birtlesii and B. grahamii, isolated from Apodemus sp. mouse and Microtus agrestis vole, respectively (Boulouis et al. 2001, Koesling et al. 2001, Marignac et al. 2010). A recent study reported a sustained bacteremia (11 weeks) in M. musculus with two B. tribocorum strains isolated from Mus caroli and Mus cervicolor, evidencing the congenic potential transmission of bartonellae (Colton and Kosoy 2012). Similarly, Swiss Webster mice inoculated with a Bartonella strain closely related to B. coopersplainensis isolated from R. norvegicus became bacteremic during a period that lasted from 4 to 8 weeks (Colton et al. 2011). Nevertheless, in the same study, bacteremia was not obtained when three other rat-associated bartonellae strains were used. Therefore, experimental evidences raised a dual scenario of what might be happening in nature: On one hand, some bartonellae may have a limited host specificity range, whereas certain spillover of other bartonellae between animals may occur. The reasons for this dichotomy must rely on the Bartonella strain and its armament of virulence factors that provide or limit its capability to “jump” from one host species to another, as described in B. grahamii (Berglund et al. 2009).
When murine challenge studies have applied to more phylogenetically distant Bartonella spp., such as B. henselae from cats, mimicking natural incidental infections, pathological manifestations were observed. It is noteworthy in these cases that bacteremia was not established (Regnath et al. 1998, Kunz et al. 2008).
Bacterial dose, required to generate an effective infection, is an important variable that should be taken into consideration in the study of Bartonella infections. In this regard, contrasting results have been obtained under experimental challenges. In one study, more mice became infected when higher doses were inoculated (Colton et al. 2011). Conversely, longer duration and higher bacteremia levels have been obtained with low inoculum doses (∼103 colony-forming units [CFU]) (Kosoy et al. 1999, Marignac et al. 2010). Interestingly, Boulouis et al. (2001) did not find significant differences in the bacteremia level with inoculum doses above 1.5×103 CFU. In host communities, rodents would likely be exposed to low doses of bartonellae by vector transmission. Notwithstanding, the number of fleas infesting each rodent and the rate of blood feeding of each flea are extra variables that can contribute to the real dose and exposure to bartonellae, as discussed before. Therefore, quantification of the bartonellae load in fleas and their gastrointestinal contents and tissues is crucial. In a pioneering study, bartonellae loads in C. felis feces were determined, reaching up to 5.3×103 B. henselae CFU/mg of feces (Finkelstein et al. 2002). More recently, through a kinetics study of B. henselae infection in cat fleas fed on an artificial feeding system, a potential replication and prolonged persistence of the Bartonella in fleas and their feces was suggested (Bouhsira et al. 2013). In addition, the determination of potential substances that could enhance the bartonellae replication or establishment of the infection in the host skin should be explored as they may reveal important features.
Field studies: Natural dynamics of bartonellae infection
To describe the temporal dynamics of Bartonella infection in rodent communities, longitudinal field studies have been carried out (Fichet-Calvet et al. 2000, Birtles et al. 2001, Kosoy et al. 2004a, Telfer et al. 2007a, 2007b, Bai et al. 2008b, Welc-Faleciak et al. 2010, Bai et al. 2011). These studies have shown interesting variations in the pattern of Bartonella infections among seasons, rodent age cohorts, rodent sexes, in dependence of the level of flea parasitism, persistence of infection, and other factors in different rodent species and communities.
As rodents' density and activity vary across seasons (Telfer et al. 2007b), the exposition and prevalence of microbial infections can also be affected by this seasonality (Bai et al. 2008b). Microbial prevalence in hosts can vary due to changes in infection resistance, association between other microbial species, and changes in the activity of their vectors (Telfer et al. 2007b). Accordingly, it has been observed that Bartonella prevalence varies significantly during the year and across years (Bai et al. 2008b, Welc-Faleciak et al. 2010). Peaks of higher Bartonella prevalence have been reported from summer to fall in most of the rodent species worldwide (Fichet-Calvet et al. 2000, Jardine et al. 2006b, Telfer et al. 2007a, Bai et al. 2008b, Paziewska et al. 2012a). Even in cotton rats (S. hispidus) from Georgia in the United States that maintain a high prevalence all over the year, a peak during early summer to mid-fall was evident (Kosoy et al. 2004a). Increasing prevalence of Bartonella infection in those warmer periods, correlates with the emerging of juvenile rodents from their burrows (Jardine et al. 2006b), higher activity of rodents, peak of the vector activity, or higher infestation by seasonal fleas (Krasnov et al. 2002, 2005). Interestingly, bank voles and wood mice from the United Kingdom demonstrated two patterns of seasonality depending on the Bartonella sp. involved (Telfer et al. 2007b). Additionally, the survival rates after a critical period (e.g., winter or summers) is a relevant variable for the Bartonella cycle, as this may remodel the host population demographics (Fichet-Calvet et al. 2000, Welc-Faleciak et al. 2010).
Age- or body mass–dependence patterns in the prevalence of Bartonella infection have been found in wild rodents. Generally, juvenile and subadult animals tend to be more affected than adults in the wild (Fichet-Calvet et al. 2000, Kosoy et al. 2004a, Jardine et al. 2006b, Telfer et al. 2007a, Bai et al. 2008b, Morway et al. 2008). This age bias has been associated with several factors, such as a tendency of juvenile rodents to be more mobile than territorial adults (higher exposure in the formers), higher risk of acquiring the infection as a result of its immature immune system, or the fact that older rodents have the chance to clear the infection during their lifetime (Fichet-Calvet et al. 2000, Welc-Faleciak et al. 2010). An acquired humoral immunity was previously proposed for the lower infection prevalence in older rodents (Fichet-Calvet et al. 2000), but low titers of antibodies observed in cotton rats have challenged this explanation (Kosoy et al. 2004a). The latter authors suggested a potential immune tolerance as an outcome of exposure to the bartonellae antigens in utero or antibody sequestration as explanation of the nondetection of antibodies, but this can be true only for those host species that present vertical transmission. Bai et al. (2008b) observed that juvenile black-tailed prairie dogs became infected shortly after emerging from their burrows, but once they reached a body mass above 300 grams, the infection prevalence declined. The authors suggested that a mass threshold (>700 grams) can represent a dramatic adjustment of their immune system linked to the clearance of the infection (Bai et al. 2008b). Furthermore, a natural selection for noninfected animals could explain the reduced prevalence of Bartonella in older animals (Jardine et al. 2006b). Nevertheless, no specific rule for Bartonella infection can be concluded, since no correlation between age groups and weight-dependent prevalence patterns were observed both in deer mice from Colorado (Bai et al. 2011) and bank voles from France (Buffet et al. 2012).
Despite sex differences in rodent ecology, contradictory results of the role of rodent sex in Bartonella infection have been observed. No difference between infection rates of males and female rodents has been observed in many studies (Fichet-Calvet et al. 2000, Kosoy et al. 2004a, Bai et al. 2008b, Morway et al. 2008, Bai et al. 2011). However, differences in the Bartonella prevalence according to sex was reported in Apodemus sylvaticus and A. flavicollis, in which males presented higher infection prevalence in comparison to female rodents (Welc-Faleciak et al. 2010, Harrison et al. 2012).
The correlation between flea abundance and bartonellae infection in rodents has shown some variation across studies. In certain cases, the seasonality of the flea positively correlated to the bartonellae prevalence in rodents (Jardine et al. 2006b, Welc-Faleciak et al. 2010). For instance, it was noticed that the higher flea abundance on wild rodents from Poland in 2006 correlated with a higher overall Bartonella infection compared to 2004 (Welc-Faleciak et al. 2010). On the contrary, differences in infestation rates of R. rattus and R. norvegicus from South Africa did not explain the observed difference in Bartonella infection prevalence (24% versus 5%) between these rat species (Brettschneider et al. 2012a). The Bartonella sp. involved can also determine the final effect of fleas on the Bartonella prevalence, as was reported on field voles (M. agrestis) in which the presence of fleas had only a positive effect on the infection probability of a Bartonella sp. (i.e., BGA strain) (Telfer et al. 2007a). Moreover, the role of fleas as intermediaries of the natural cycle was proposed for B. taylorii in wood mice (A. sylvaticus) and B. doshiae in field voles (M. agrestis), since a “delayed effect” of host densities was noticed (i.e., fleas buffering the timing of transmission) (Telfer et al. 2007a, 2007b).
Variations in the persistence of Bartonella infection in wild conditions have also been observed. Some studies on rodent populations from Europe and United States have reported long-persistent bacteremia of several months (Kosoy et al. 2004a, Bai et al. 2011). Through sequential screenings of tagged individual rodents, the Bartonella infection has shown to be very dynamic. Some rodents demonstrated a clearance of the infection in a short time (Birtles et al. 2001), whereas others remained infected for periods as long as 9 months (Kosoy et al. 2004a). In addition, reappearance of bacteremia after an apparent infection clearance has also been observed in wild rodents (Kosoy et al. 2004b, Jardine et al. 2006a, Bai et al. 2011).
Many rodent populations exhibit certain features that may play a role in the Bartonella prevalence and distribution. The northern grasshopper mouse (O. leucogaster) routinely invades burrows of other rodents and predates on their owners (McCarty 1978). This behavior can be associated with the remarkable prevalence of Bartonella infection in O. leucogaster, one of the highest detected to date (Bai et al. 2007a). On the contrary, there are cases in which ecological characteristics can hardly be associated with the Bartonella infection. Two closely related Neotoma spp. demonstrated a different Bartonella prevalence, despite the fact that their habitat, diet, behavior, and parasitizing flea species are virtually the same (Morway et al. 2008). Similarly, the difference in the Bartonella persistence on M. glareolus and A. flavicollis seems to lie in the bacteria–rodent interaction rather than ecological differences between these rodents (Paziewska et al. 2012a).
Bartonella Infection Composition
Infection with more than one Bartonella spp. or variant is a well-acknowledged phenomenon in rodents and fleas (Kosoy et al. 2004b, Abbot et al. 2007, Telfer et al. 2007a, Brinkerhoff et al. 2010, Morick et al. 2011). The determination of the infection composition in the host and vectors can help understanding the infection fluctuations observed in longitudinal studies. Kosoy et al (2004b) isolated up to three different genotypes from a single cotton rat blood sample. The authors detected co-infection in 21.3% of 408 cotton rat samples holding an overall of 26 combinations of mixed strain infections. In a recent study, we observed that multiple variants can be distributed in rodents and their fleas in an expanded repertoire of infection compositions (Gutiérrez et al. 2014). Single carriers (rodent and fleas) harbored infection composed of multiple closely and distantly phylogenetically related Bartonella genotypes, which circulated under a potential intergenotype competition and reflected a tendency to dominate a particular carrier type (i.e., rodent or flea). It seems that traditional diagnostic methods could bias the real picture of Bartonella infection structures in reservoir animals.
Longitudinal studies have shown that in sequential screenings a Bartonella genotype can be repetitively detected or replaced by a close or distinct phylogenetic genotype, even after a nonbacteremic period (Birtles et al. 2001, Kosoy et al. 2004b, Jardine et al. 2006a). New bacteremic periods of a previously detected genotype could be the result of a new vector transmission event or a reseeding event of the hidden genotype from the potential primary niche (as previously discussed) (Schulein et al. 2001, Kosoy et al. 2004b, Bai et al. 2011). An alternative explanation might be that the infection has never been cleared from the blood, but its level decreased below detectable levels (Bai et al. 2011). Thus, this fluctuation in Bartonella in wild rodents (alternating bacteremic and nonbacteremic periods) has questioned the effectiveness of the host immune system.
Co-infection with more than one Bartonella species in wild rodents might reveal an additional scenario associated with the cyclic bacteremia phenomenon. In a previous study, two different Bartonella strains were isolated at different sampling dates from the blood of naturally infected rodents (M. crassus and Gerbillus nanus) kept under laboratory conditions (i.e., no external source of infection) (Morick et al. 2011). Similarly, different B. henselae variants were isolated from different bacteremic peaks in naturally infected cats (Kabeya et al. 2002). These findings suggest that two co-existing Bartonella variants could alternate between the primary niche and the bloodstream, producing individual bacteremic waves. Another option is that both variants can co-exist in the blood, but one dominates the infection and can obscure the presence or detection of other variants, as observed in the Bartonella infection composition in wild rodents and their fleas (Gutiérrez et al. 2014). Nevertheless, Chan and Kosoy (2010) explored the frequencies of acquiring new variants in the cotton rats, and concluded that some cross-immunity exists and plays a filtering role, limiting the reinfection of some variants that are closely related. It seems evident that in sequential determinations either the different variant infections are intercalating in a way that a long persistent infection is maintained (yet heterogeneous), or a multi-infection with different variants is occurring more frequently than expected, and the determination of the predominant variant in a given time obscures the others (Paziewska et al. 2012a).
Conclusions
Bartonellae have shown an outstanding adaptation within rodents and their flea parasites. The diversity of Bartonella spp. and genotypes discovered in Rodentia compared to other mammals is the highest to date. Additionally, spillover of bartonellae between rodent species and genera, and the interactions between co-existing Bartonella variants in the same host, might play a substantial role in the generation of diversity in these bacteria. Investigation of the ecology of populations and communities of rodents and fleas can help understand the dynamics of Bartonella infections, but generalizations are not apparent across rodent–flea systems. Much contrasting evidence has raised a puzzle of ecological traits affecting the Bartonella cycle between the rodent populations studied to date. It appears that the interactions within the bartonellae–flea–rodent triangle are specific and result in particular traits for each system. Thus, the elucidation of each Bartonella cycle must be evaluated individually.
Footnotes
Acknowledgments
This research was supported by the Israel Science Foundation (grant number 30/11 to Shimon Harrus), the Ministerio de Ciencia y Tecnologia (MICIT), and the Consejo Nacional para Investigaciones Cientificas y Tecnologicas (CONICIT), Costa Rica. This is publication no. 835 of the Mitrani Department of Desert Ecology.
Author Disclosure Statement
No competing financial interests exist.