
Abstract
Since 2002, an increased number of northern sea otters (Enhydra lutris kenyoni) from southcentral Alaska have been reported to be dying due to endocarditis and/or septicemia with infection by Streptococcus infantarius subsp. coli. Bartonella spp. DNA was also detected in northern sea otters as part of mortality investigations during this unusual mortality event (UME) in Kachemak Bay, Alaska. To evaluate the extent of exposure to Bartonella spp. in sea otters, sera collected from necropsied and live-captured northern sea otters, as well as necropsied southern sea otters (Enhydra lutris nereis) unaffected by the UME, were analyzed using an immunofluorescent antibody assay. Antibodies against Bartonella spp. were detected in sera from 50% of necropsied and 34% of presumed healthy, live-captured northern sea otters and in 16% of necropsied southern sea otters. The majority of sea otters with reactive sera were seropositive for B. washoensis, with antibody titers ranging from 1:64 to 1:256. Bartonella spp. antibodies were especially common in adult northern sea otters, both free-living (49%) and necropsied (62%). Adult stranded northern sea otters that died from infectious causes, such as opportunistic bacterial infections, were 27 times more likely to be Bartonella seropositive than adult stranded northern sea otters that died from noninfectious causes (p<0.001; 95% confidence interval 2.62–269.4). Because Bartonella spp. antibodies were detected in necropsied northern sea otters from southcentral (44%) and southwestern (86%) stocks of Alaska, as well as in necropsied southern sea otters (16%) in southcentral California, we concluded that Bartonella spp. exposure is widely distributed among sea otter populations in the Eastern Pacific, providing context for investigating future disease outbreaks and monitoring of Bartonella infections for sea otter management and conservation.
Introduction
N
Because a large number of northern sea otters were found dead in southcentral Alaska beginning in 2006, the US Working Group on Marine Mammal Unusual Mortality Events declared an unusual mortality event (UME; Gill 2006). Since that time, diagnostic efforts have been directed toward understanding the causes of the sea otter UME. Streptococcus infantarius subsp. coli (a member of Streptococcus bovis/equinus complex, SB/E) infection was identified to be highly associated with the mortality in Kachemak Bay and surrounding areas. This bacterium was isolated from heart valves of northern sea otters with vegetative valvular endocarditis (VVE), with or without septicemia (Gill 2006). Recently, we reported that up to 45% of examined northern sea otters had detectable Bartonella spp. DNA, and 33% of the heart valves from animals that died with VVE were co-infected with Bartonella spp. and S. infantarius subsp. coli (Carrasco et al. 2014).
Bartonella spp. are fastidious Gram-negative bacteria that infect erythrocytes and vascular endothelial cells of hosts and are usually transmitted by blood-sucking arthropods (Chomel et al. 2009). This facultative intracellular bacterium causes persistent asymptomatic bacteremia and has been associated with debilitating and possibly life-threatening illnesses, including encephalopathies, bacillary angiomatosis, myocarditis, and valvular endocarditis in domestic animals and humans (Breitschwerdt et al. 2010). Clinical manifestations vary on the basis of the immune status of the host, the infecting Bartonella strain, and the co-evolutionary history of a Bartonella spp. with its animal host (Chomel et al. 2009). In their mammalian reservoir hosts, Bartonella spp. often cause asymptomatic chronic bacteremia, whereas in the nonreservoir hosts this pathogen is typically associated with multiple disease processes (Chomel et al. 2009). Bartonella spp. have been increasingly isolated or detected in blood or lesions from live-captured or moribund marine animals (Maggi et al. 2008, Morick et al. 2009); however, little is known about the epidemiological significance, as hosts and transmission dynamics of this organism in the marine environment are relatively unstudied.
Because Bartonella DNA was recently detected in northern and southern sea otter carcasses from Alaska and California (Carrasco et al. 2014), our main objective was to assess exposure in northern and southern sea otters to this usually vector-borne pathogen. The goals of this study were two-fold: (1) To determine the frequency and distribution of serum antibodies against selected Bartonella species in northern and southern sea otters and (2) to evaluate risk factors associated with Bartonella exposure in both populations. Our findings provided baseline serological data and insight into the epidemiology of bartonellosis in sea otters to contribute to conservation efforts.
Materials and Methods
Sampling
Serum samples were collected from 44 apparently healthy, live-captured northern sea otters sampled in Kachemak Bay, Alaska, in 2007 by the United States Fish and Wildlife Service (USFWS), Anchorage, Alaska. Blood was collected from anesthetized sea otters using well-established protocols for immobilization, sampling, and release (Doroff and Badajos 2010). Samples were also obtained from 48 northern sea otters that stranded and were necropsied between 2004 and 2009. Postmortem examinations of northern sea otters were performed at the Anchorage USFWS office by a veterinary pathologist or by veterinarians with marine mammal necropsy experience.
At necropsy, northern sea otters were categorized into either the infectious or noninfectious group on the basis of the diagnosed cause of death, such as opportunistic bacterial infections. Thirty-three individuals were categorized into an “infectious” group, whereas those animals dying due to trauma and/or other causes and with no evidence of concurrent infectious disease were included in a “noninfectious” group (n=15). Bacterial pathogens isolated or detected in lesions of necropsied northern sea otters included S. infantarius subsp. coli or SB/E organisms, Bartonella spp., and Streptococcus phocae, as previously described (Carrasco et al. 2014). Stranded sea otters were found dead or moribund in south-central Alaska, Kodiak Archipelago, and the western side of Cook Inlet, including Ninilchik, Homer and Kachemak Bay, Whittier, Kodiak Island, Hallo Bay, and Kamishak Bay (Fig. 1). Sea otters from southcentral stock that had stranded in the Ninilchik, Whittier, Homer, and Kachemak Bay regions were further subcategorized into a “Kachemak Bay area” group, whereas animals from southwest stock that had stranded in Kamishak Bay, Hallo Bay, and Kodiak Island were pooled into a “southwest Alaska” group. These categories were selected because mortalities caused by SB/E infection were common in northern sea otters from southcentral stock in Kachemak Bay and surrounding areas (adjacent to the threatened southwestern stock).
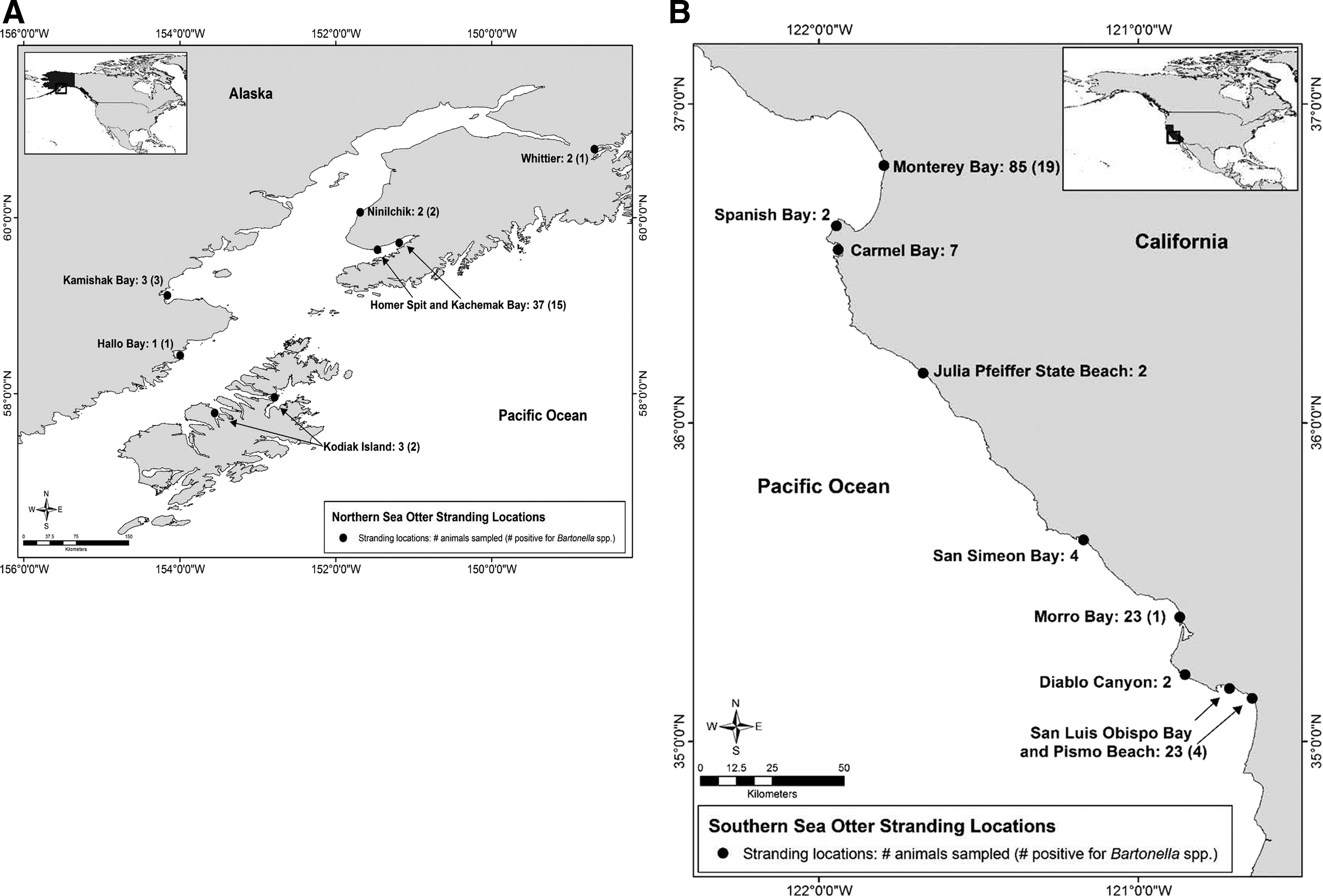
Maps of Alaska and California showing stranding areas of northern and southern sea otters that were necropsied and tested by indirect immunofluorescent antibody (IFA) assay for Bartonella spp. (
In addition, 148 serum samples were obtained from necropsied southern sea otters that had stranded during 2001–2009 in California. Postmortem examinations of southern sea otters were performed at the Marine Wildlife Veterinary Care and Research Center, California Department of Fish and Wildlife (CDFW), Santa Cruz, California, by a veterinary pathologist to establish the cause of death. As with the northern sea otters, southern sea otters that were classified into the “infectious” group generally were diagnosed with a cause of death due to either bacterial or parasitic pathogens or had bacterial or parasitic infections as a contributing cause of death (n=72 animals), whereas animals that were diagnosed with other noninfectious causes of death or had contributing factors that were not associated with infectious disease were included in a “noninfectious” group (n=76). Acknowledged pathogens of southern sea otters included a range of bacterial, protozoal, and acanthocephalan parasites, as previously described (Kreuder et al. 2003, Miller et al. 2010a). These sea otters were found dead or moribund on beaches in various geographic areas along the central California coast, including Monterey Bay, Spanish Bay, Carmel Bay, Julia Pfeiffer State Beach, San Simeon Bay and surroundings, Morro Bay and surroundings, Diablo Canyon, and San Luis Obispo Bay and Pismo Beach (Fig. 1.).
The age of each sea otter was estimated at the time of necropsy based on body mass, weight, dentition, pelage characteristics, and reproductive status (Riedman and Estes 1990, Von Biela et al. 2011). Pups, juveniles, and subadults were categorized as “immature” otters, and adults and aged adults were categorized as “adult” otters. Nutritional condition was assessed by the amount of subcutaneous fat present and by palpation of the ribs, backbone, and other bony protuberances (Doroff and Mulcahy 1997). Otters were categorized as being in “poor” nutritional condition if they were thin or emaciated with scant subcutaneous fat and evidence of muscle loss, or in “good” nutritional condition if they had moderate to abundant subcutaneous fat.
Serology
Antibodies against Bartonella henselae type II (also known as Marseille; isolate U4, University of California–Davis), Baronella clarridgeiae (ATCC 51734), and Bartonella washoensis (isolate UCD-dog2, University of California–Davis) in serum and/or whole blood samples from northern and southern sea otters were detected using an indirect immunofluorescent antibody (IFA) assay. The IFA procedure was similar to the procedure described by Henn et al. (2007) with a slight modification. Fluorescein isothiocyanate (FITC)-conjugated goat anti-ferret immunoglobulin G (IgG) (Bethyl Laboratories, Montgomery, TX) was used for detection of primary antibodies. B. henselae, B. clarridgeiae, and B. washoensis strains were used as surrogate bacteria for this assay due to the lack of success obtaining a Bartonella isolate from sea otter tissues. Samples were examined at dilutions of 1:64 and 1:128 using a fluorescence microscope at 400×magnification. The intensity of bacillus-specific fluorescence was scored subjectively from 1 to 4, as previously described (Chang et al. 2000). Samples with a fluorescence score of ≥2 at a dilution of 1:64 were considered positive because this antibody titer has proven to indicate prior exposure and current infection by Bartonella spp. in dogs and cats (Chomel et al. 1995, Pappalardo et al. 1997). Samples testing positive were titrated in serial two-fold dilutions until the fluorescence score was below the limit of detection (<2). Each slide was examined in a blinded fashion by three readers, and fluorescence scores were averaged.
Statistical analysis
Data collected on each necropsied northern and southern sea otter included animal identification number, age class, sex, stranding date and location, date of sampling, weight, nutritional condition, and cause of death from necropsy findings, including pathological lesions and reported laboratory results for any postmortem bacterial culture or parasitology result. Additional SB/E or S. infantarius subsp. coli and S. phocae culture results were provided by the USFWS from microbiological reports from the Microbiology Laboratory (Veterinary Medical Teaching Hospital, University of California–Davis). Chi-squared analyses were used to evaluate associations between Bartonella IFA results and age, sex, nutritional condition, stranding area, and animal grouping as “infectious” or “noninfectious” cause of death using JMPv8.0 software (SAS Institute Inc,
Results
Serological findings
Antibodies reactive to Bartonella spp. were detected in 34% (15/44) and 50% (24/48) of serum samples from live-captured and necropsied northern sea otters, respectively, with antibody titers ranging from 1:64 to 1:256 (Tables 1 and 2). By comparison, Bartonella spp. seroprevalence was lower (16%, 24/148) for southern sea otters (Table 1). The majority of positive sea otter sera were reactive for B. washoensis, as antibodies reactive to B. washoensis (≥1:64 IFA titers) were detected in 27% (12/44) of sera from live-captured northern sea otters (Table 1). Two (4.5%, 2/44) were seropositive for B. clarridgeiae and B. washoensis, and one sample was seropositive only to B. clarridgeiae (≥1:128). Of 48 necropsied northern sea otters sampled, seven (14%, 7/48) reacted only to B. washoensis and 12 (25%, 12/48) were reactive to B. henselae and B. washoensis (Table 1). Antibodies reactive to B. washoensis (≥1:64) were detected in 13% (19/148) of sera from necropsied southern sea otters (Table 1). Seven samples (4.7%, 7/148) were seropositive only to B. washoensis, and eight samples (5.4%, 8/148) tested positive for B. henselae and B. washoensis.
ND, Not detected.
Antibody titers ≥1:64 were classified as seropositive.
IFA, indirect immunofluorescent assay.
Association of Bartonella seroprevalence with demographic and spatial factors
Significant associations were identified between Bartonella seropositivity in northern sea otter carcasses and adult age class, poor nutritional condition, and death due to infectious causes (Table 3). Bartonella seropositivity was not more common in necropsied northern sea otters when compared to apparently healthy live animals. A much higher proportion of Bartonella-seropositive stranded northern sea otters died with infectious disease as a primary or major contributing cause of death, when compared to stranded northern sea otters that died from noninfectious causes (Table 3). Adult and aged adult stranded northern sea otters that died with infectious disease were 27 times more likely (Fisher exact test, p<0.001; 95% CI 2.62–269.4) to be seropositive to Bartonella spp. than northern sea otters that died from noninfectious causes. A similar age association was observed for Bartonella-seropositive, live-captured northern sea otters (p= 0.04). Of 28 adult, live-captured northern sea otters tested, 12 (49%) were seropositive, compared to two out of 16 immature sea otters (13%; p= 0.04).
In contrast to observed patterns for northern sea otters, no significant associations were observed between Bartonella seropositivity and age class, sex, nutritional condition, or cause of death for southern sea otter carcasses. Southern and northern sea otter carcasses that tested positive for Bartonella spp. were found on beaches in various geographic areas of California and Alaska (Fig. 1).
Discussion
The high proportion of B. washoensis–seropositive sea otters was consistent with our recent detection of Bartonella DNA in sea otter heart valves from Alaska and California (Carrasco et al. 2014). The Bartonella DNA detected was closely related to B. henselae, B. washoensis, and Candidatus Bartonella volans, suggesting that northern and southern sea otters are infected with more than one Bartonella species (Carrasco et al. 2014). Consistent with that finding, seroreactivity to greater than one Bartonella spp. was detected in 67% (16/24) and 50% (12/24) of B. washoensis–positive samples from northern and southern sea otter carcasses, respectively. This result was not surprising given that serological cross-reactivity of these three Bartonella spp. and others has been previously reported in sera from Bartonella-infected humans and animals (Maurin et al. 2002, Chomel et al. 2003, Tsuneoka et al. 2004, Boulouis et al. 2005). In contrast to samples from sea otter carcasses, the majority of the sera from presumably healthy, live-captured northern sea otters showed exclusive reactivity against B. washoensis (86%; 12/14 B. washoensis–positive samples). The combined PCR (Carrasco et al. 2014) and serology results from this study indicated that at least B. washoensis–like organisms were circulating among northern sea otters, consistent with recent findings for closely related American river otters (Lontra canadensis), that were also infected with either B. washoensis–like or B. volans–like organisms (Chinnadurai et al. 2010).
Bartonella exposure was detected among all northern sea otter age groups, but it was most common in adults. This finding is consistent with serosurveys of B. henselae in cats (Guptill et al. 2004) and Bartonella vinsonii subsp. berkhoffii in coyotes (Chang et al. 1999, Chang 2000) that showed that Bartonella seroprevalence increased with age, whereas Bartonella bacteremia decreased as the age of these other species increased. Because seropositivity was common and the risk of exposure increased with age, it is likely that Bartonella spp. exposure is endemic in northern sea otters in Alaska. Bartonella spp. are usually transmitted among terrestrial reservoirs by blood-sucking arthropods such as sand flies, fleas, or lice (Chomel et al. 2009); however, little is known regarding potential vectors for Bartonella spp. transmission in aquatic animals. Interestingly, Bartonella DNA identical to that from Bartonella-infected harbor seals was detected in hematophagous lice that are common pinniped ectoparasites (Thompson et al. 1998, Morick et al. 2009). This finding suggested that marine host-adapted vectors may be able to transmit Bartonella spp.
Reports of co-infections of Bartonella spp. and other opportunistic pathogens are common in domestic animals and humans (Breitschwerdt and Kordick 2000, Boulouis et al. 2005). Our investigation revealed a similar relationship between Bartonella seropositivity and death due to infectious causes for stranded northern sea otters, as 16 of the 22 seropositive animals from the infectious group died with SB/E-associated endocarditis or septicemia. Furthermore six of these 22 seropositive northern sea otters were co-infected with Bartonella DNA and SB/E organisms, as was recently described (Carrasco et al. 2014). Interestingly, five of these six seropositive animals had antibodies reactive to B. washoensis and detectable Bartonella DNA that was closely related to B. washoensis and Candidatus B. volans (Carrasco et al. 2014). Some B. washoensis–seropositive northern sea otters that had died due to VVE yielded titers ≥1:256, as commonly reported in Bartonella-associated endocarditis in dogs and humans (Maurin et al. 2002, MacDonald et al. 2004). However, the clinical impact of disseminated Bartonella infection in sea otters was not clear. The observed high mortality due to VVE for northern sea otters from this region and confirmation of co-infection by Bartonella and other pathogenic bacteria in prior studies (Brownstein et al. 2011, Carrasco et al. 2014) suggests that Bartonella co-infections may be a contributor to the development of debilitating disease in northern sea otters.
Previous studies in dogs and humans with infective endocarditis have shown evidence of serological cross-reactivity between Bartonella spp. and intracellular bacteria (LaScola and Raoult 1996, Maurin et al. 2002, Macdonald et al. 2004). Although it is reasonable to speculate that SB/E-infected northern sea otters could have cross-reactive antibodies to Bartonella antigens using our IFA-based assay, data from our laboratory and others have shown that cross-reactivity between Bartonella spp. and Streptoccocus spp. is rare when samples are evaluated with this serological assay (Sykes et al. 2006, Vermeulen et al. 2010). However, a potential limitation of our findings is that we cannot rule out the possibility of cross-reactive antibodies to other intracellular pathogens that have shown to interfere in the diagnosis of bartonellosis in dogs and humans (Maurin et al. 2002, MacDonald et al. 2004).
Exposure to Bartonella spp. was widely distributed among sea otters from the southcentral stock of Alaska, because positive samples were detected in otters from Ninilchik, Whittier, and Kachemak Bay. In addition, Bartonella spp. antibodies were detected in the southwestern stock of northern sea otters that inhabit areas adjacent to Kachemak Bay, including Kamishak Bay and Kodiak Island. This broad distribution of seropositivity suggested that this bacterium is endemic in northern sea otters. Due to shared haul-out sites and rafting behavior of sea otters in southcentral Alaska (Doroff and Badajos 2010) and the wide range and diversity of seroprevalence to Bartonella spp. documented here, it appears that Bartonella may be transmitted horizontally between sea otters, possibly by yet unidentified arthropod vector/vectors. It is also possible that a common vector may be involved in the transmission of Bartonella between northern sea otters and river otters since fecal analyses in haul-out sites in Kachemak Bay showed that otters likely used the same haul-out sites as coastal river otters at different times of day (Doroff et al. 2012).
In conclusion, this study together with our recently published molecular findings (Carrasco et al. 2014) confirms that Bartonella organisms have been present in sea otters over the past decade and provides evidence for widespread exposure to Bartonella spp. among northern and southern sea otters. This bacterium seems to be regionally endemic for northern sea otter subspecies in Kachemak Bay and surrounding areas. Our findings provide baseline data on the frequency and distribution of Bartonella exposure among northern and southern sea otters in Alaska and California, providing context for investigating future disease outbreaks and monitoring of Bartonella infections for sea otter management and conservation.
Footnotes
Acknowledgments
The authors thank the personnel from the US Fish and Wildlife Service (USFWS), Anchorage, Alaska; Alaska SeaLife Center, Seward, Alaska; and the Marine Wildlife Veterinary Care and Research Center, California Department of Fish and Wildlife (CDFW), Santa Cruz, California; as well as the Homer stranding network for assistance with animal captures, necropsies, and sample collection. We also thank Kate Thomas for helping to illustrate sampling areas and Dana Jenski (USFWS) and Erin Dodd and Eva Berberich (CDFW) for their assistance on demographic and spatial information of sea otters and logistics for collection and shipping samples. Field sample collections in Alaska were conducted under authority of a Marine Mammal Protection Act permit issued by the Division of Management Authority, USFWS (MA041309-4 permit number) and under an Insitutional Animal Care and Use Committee (IACUC) review by the University of Alaska. This research was supported by grant no. D08ZO-046 from the Morris Animal Foundation; the Karen C. Drayer Wildlife Health Center graduate student fellowship, University of California, Davis; and an Oceans and Human Health training grant no. NA06OAR4310119 (Training Tomorrow's Ecosystem and Public Health Leaders Using Marine Mammals as Sentinels of Oceanic Change) from the National Oceanic and Atmospheric Administration (NOAA). S.E.C. was also supported by grant no. NA06OAR4310119 and National Institutes of Health National Research Service Award T32 AI 060519.
Author Disclosure Statement
No competing financial interests exist.
The findings and conclusions in this article are those of the authors and do not necessarily represent the views of the US Fish and Wildlife Service.